

22 Gen 30. Interazioni tra organismo e ambiente e tra distretti dell’organismo. Le basi fisiopatologiche delle Terapie con Anestetici Locali
Claudio Dell’Anna, Massimo Barrella
E’ sufficiente spiegare il rationale della Terapia Neurale secondo Huneke (TNH) semplicemente invocando la connettività del Sistema Nervoso Vegetativo (SNV) che permette a un focolaio irritativo periferico di manifestare i suoi effetti nocivi anche a distanza dalla sua origine? Quasi certamente no: la spiegazione richiederebbe l’illustrazione di elementi connessi da relazioni complesse (non-lineari) mentre siamo incessantemente bombardati da formulazioni scientifiche in cui la linearità dei nessi causali è proposta in forma artificiosamente semplificata.
L’avvento del microscopio determinò il tramonto di numerose convinzioni metafisiche, le malattie contagiose cessarono di essere viste quali effetti di influssi maligni e l’idea che le guarigioni si dovessero al fatto che le preghiere avessero raggiunto il cielo perse popolarità. Tutto questo però non condusse la Medicina a un punto fermo, non si pervenne a certezze assolute. Più di cento anni sono trascorsi dall’inizio della contrapposizione tra le idee partorite da due “prìncipi del microscopio”, Louis Pasteur (1822-1895) e Pierre Jacques Antoine Béchamp (1816-1908). Il primo sostenne il principio del “monomorfismo”: le malattie si generano per aggressione dall’esterno di microrganismi che nel passaggio da un ospite all’altro mantengono la loro identità inducendo sempre la stessa forma morbosa secondo il principio “un batterio, una forma, una malattia”. Béchamp difese invece l’idea del “pleiomorfismo”: un microrganismo proveniente dall’ambiente esterno può esibire diversi gradi di patogenicità in rapporto alle condizioni dell’ospite entro cui può generare numerose condizioni (francamente morbose e non) variando la sua forma originaria secondo il principio “un batterio, più forme, più malattie”. Al netto delle declinazioni più esasperate che le due tesi hanno espresso nella comunità scientifica è innegabile che la seconda sia stata supportata da un maggior numero di riscontri. In ogni caso ancora oggi entrambe coesistono dato che da un lato impieghiamo di antibiotici e dall’altro cerchiamo protezione dalle infezioni osservando salutari stili di vita. Si può dire che ognuna delle due abbia generato un proprio “mercato”, abbia formulato una propria filosofia. Ci troviamo forse di fronte a “due Medicine”? Certamente no: si tratta di due punti di vista ognuno dei quali offre il proprio apporto ed è innegabile che l’umanità abbia tratto vantaggi da entrambi. Seppure con argomentazioni diverse entrambi hanno avuto il merito di presentare il percorso biologico dei viventi quale risultato di incessanti interazioni tra organismo e ambiente e tra diversi settori di ogni organismo. Perciò è importante anzitutto comprendere almeno per sommi capi le leggi a cui quelle interazioni sono assoggettate e le dinamiche con cui possono sottrarsene. In questa luce assume il valore di pietra angolare l’opera di Gustav Ricker dedicata alla teoria da lui formulata della “Patologia di relazione”: vent’anni prima della nascita della TNH lo studioso di Hadamar aveva dimostrato sperimentalmente la relazione tra settori somatici lontani inducendo fenomeni vasomotori diffusi all’intero organismo a seguito di stimoli puntuali [43]. Quanto detto fin qui basterebbe a sollecitare la riflessione sui danni che la Medicina soffrirebbe per un “pensiero unico” imposto da qualunque entità che abbia la forza di farlo: si tornerebbe in una dimensione prescientifica in cui l’idea diversa da quella dominante assumerebbe il significato di eresia divenendo oggetto di marginalizzazione. Quella che viene indicata con il termine onnicomprensivo di “Medicina” è in realtà una miscela di attività tra loro anche assai diverse sostenute da altrettanto diverse forme di pensiero: la Chirurgia si fonda sull’idea di ripristinare la salute modificando la struttura materiale degli organi, l’Antibioticoterapia si basa sulla vulnerabilità dei microrganismi a sostanze studiate al fine di combatterli, la Fisioterapia valuta i difetti funzionali e applica strategie per il recupero delle funzioni perdute. Si potrebbero ancora citare altre forme di Medicina ma, per quanto diverse nella prassi e nel retrostante pensiero, tutte possono efficacemente combinarsi nella finalità del ripristino dello stato di salute: in caso di chirurgia protesica articolare non stupisce che entro un breve arco di tempo il paziente sia sottoposto all’atto chirurgico, a una copertura antibiotica e alla fisioterapia.
Le “Terapie Funzionali”, tra cui spiccano le Terapie Neurali, mirano alla realizzazione di profili di regolazione biologica più idonei al recupero dello stato di salute e anch’esse, oltre ad aver dimostrato efficacia e versatilità in clinica, sono ben compatibili con altre forme di intervento terapeutico. Rispetto ad altre forme di Medicina si sconta però minore chiarezza sul background fisiopatologico. Occorre ammettere che il pensiero sottostante è più complesso: piuttosto che guardare allo stimolo e alla sua nocività la malattia è qui interpretata quale effetto di risposte biologiche agli stimoli non adeguate per orientamento, scansione temporale, distribuzione spaziale e ampiezza. Inoltre esse contemplano che strutture biologiche sollecitate o deteriorate possano realizzare in proprio numerosi e vari stati morbosi utilizzando le stesse dinamiche e la stessa “attrezzatura biologica” alla base del mantenimento dello stato di salute. L’analisi delle interazioni tra organismo e ambiente e tra diversi settori dell’organismo si costituisce perciò quale nucleo della cultura fisiopatologica di cui il medico, e non solo chi applichi le terapie neurali, dovrebbe disporre.
Alla ricerca di sempre maggiore chiarezza in questa direzione, nel corso della nostra quarantennale pratica di Terapie con Anestetici Locali (TAL) abbiamo consultato instancabilmente la letteratura medica internazionale e cercato occasioni di confronto. Non soltanto con i Colleghi professanti la nostra stessa attività abbiamo perseguito il confronto ma anche con interlocutori dotati di diverse competenze (scelta questa che ci ha messo in relazione con medici che delle TAL quasi neppure conoscevano l’esistenza), con fisici, chimici, biochimici e bioingegneri. Quell’orientamento antico e mai rinnegato, comprovato dalle nostre numerose pubblicazioni in convegni e riviste di diverse specialità della Medicina, si è fondato su due profonde convinzioni: anzitutto se è vero che si tratta di una metodica che merita a pieno titolo di comparire tra le opzioni di ogni medico clinico non può giustificarsi alcuna riluttanza al confronto con l’intera comunità scientifica. In secondo luogo la dimensione “di nicchia” in cui talora questa straordinaria metodica è stata compressa forse ha privato i professanti di molti stimoli che scaturiscono da ogni sano contraddittorio. Invece il confronto, persino aspro, porta del bene anzitutto favorendo il riconoscimento di alcune importanti proprietà dei principali biosistemi regolatori e proponendo una nuova formulazione di categorie già note e convalidate dal consesso scientifico. Per esplicitare tali aspetti di “novità” è stata necessaria l’adozione di alcuni termini nuovi ed è perciò che a queste righe segue un glossario. Il Lettore infatti si imbatterà frequentemente in termini/concetti inediti quali “flusso”, “leva funzionale”, “dosso” e ancora altri ma confidiamo che dopo le prime pagine potrà coglierne appieno il significato e farli propri. La comprensione di questi elementi corrobora le basi scientifiche della TNH e, più in generale, agevola il medico clinico nei passaggi dal ragionamento fisiopatologico alle scelte pratiche. Infatti a guidare la mano di chi professa le Terapie Neurali dev’essere il pensiero fisiopatologico e tanto più questo godrà di ampio respiro tanto più l’identità professionale dell’operatore potrà elevarsi. Alcuni argomenti di carattere istopatologico, pur importanti per la completezza della trattazione, non hanno potuto trovare posto qui e se ne suggerisce la lettura nel capitolo (31). Segue il glossario:
- Accentuazione/riduzione del solco mnesico: fenomeno informazionale per cui la reiterazione di una noxa su un settore tessutale produce un complesso informazionale integrato che a distanza di tempo può mediare una risposta la cui entità non è proporzionale alla noxa iniziale. L’esempio clinico più noto è la tachifilassi allergica, ma qui estendiamo il significato funzionale della definizione a tutti i processi fisiopatologici.
- Bioritmo: oscillazione nel tempo di fenomeni regolatori mediati da leve funzionali. E’ frutto dell'interazione tra due o più flussi informazionali convergenti su un punto.
- Bioritmo autogeno: reiterazione di un bioritmo connessa alla riproposizione ciclica delle leve funzionali che lo sostengono.
- Complesso informazionale integrato: insieme di elementi composti da una interazione programmata (geneticamente determinata) tra eventi biomolecolari e/o elettrici, in grado di esercitare una funzione modulatoria rispetto alla liberazione, nel tempo e nello spazio, di leve funzionali. Può essere costituita da interazioni prevalentemente biochimiche (come i complessi formati tra immunoglobuline e recettori di cellule che producono citochine) piuttosto che da interazioni prevalentemente elettrochimiche (come i neurotrasmettitori che legano recettori pre-sinaptici dei neuroni).
- Coordinazione-inibizione: intervento di istanze che modulano in senso negativo (inibizione) l’ascesa di perturbazioni moltiplicative (eccitazione) lungo un flusso informazionale longitudinale. Queste istanze si estrinsecano nella stessa disposizione geometrica tridimensionale del flusso e, al contempo, nell’intervento di azioni esercitate da complessi informazionali integrati (ormonali, immunitari, nervosi).
- Descrizione elettrica di un evento: trasformazione di una perturbazione espressa attraverso una leva funzionale biochimica, quindi analogicamente quantificabile, in una modulazione di eventi elettrici digitalizzati. Qui viene usato come formula esplicativa del fenomeno squisitamente nervoso della produzione del firing di potenziali d’azione, anche se fenomeni del tutto analoghi si ravvedono anche, ad esempio, nella trasmissione dei segnali intracitoplasmatici mediati da cariche ioniche.
- Dominio spazio-temporale: definisce l'estensione spaziale di un bioritmo. Ogni dominio spazio-temporale è portatore di un certo grado di risoluzione spazio-temporale, cioè la misura del suo rapporto tra discriminazione e reticolarità. La risoluzione spaziale tende a variare sempre insieme a quella temporale, parallelismo in cui si ravvede la fondamentale istanza “omeostatica” dei sistemi viventi.
- Flusso di informazione biochimico/nutrizionale: passaggio di istanze costituenti leve funzionali all’interno di un tragitto sequenziale che scorre disciolto in un mezzo più o meno liquido.
- Flusso di informazione contrattile e/o secretivo: passaggio di istanze costituenti leve funzionali all’interno di un tragitto sequenziale che scorre attraverso fenomeni di accoppiamento elettrochimico.
- Flusso di informazione nervoso: passaggio di istanze costituenti leve funzionali all’interno di un tragitto sequenziale che scorre attraverso la propagazione di un firing elettrico di potenziali d’azione.
- Integrazione-elaborazione: costituzione di un bioritmo, frutto dell’interazione tra flussi informazionali, assumente un grado più alto di dominio spazio-temporale per la costituzione di leve funzionali a più alto grado di simbolizzazione.
- Interprete: qualsiasi elemento in grado di accogliere un contesto biologico e trasdurlo in un codice informazionale. Questo ruolo funzionale può essere assunto da ogni tipo di elemento posto nell’interfaccia tra flussi di informazione venendosi a identificare nella recettorialità dei tessuti che delimitano i flussi stessi. Qui lo abbiamo identificato nella cellula endoteliale così come nell’eme dell’emoglobina, oltre che, naturalmente, nei recettori sinaptici dei neuroni.
- Leva funzionale: qualsiasi agente chimico-fisico in grado di esercitare il ruolo di tramite tra eventi posti in due adiacenti dimensioni spazio-temporali.
- Mappa reticolare di soglie: disposizione dei solchi mnesici nel dominio dello spazio di un organismo.
- Modalità simbolica/simbolizzazione: prodotto del fenomeno di integrazione/elaborazione consistente nella propagazione di un flusso di informazione nervoso in esito alla sottrazione del contenuto analogico da uno o più flussi di informazione posti nei domini sottostanti (biochimico/contrattile/secretivo).
- Modulazione dell'eccitabilità: risultato dell’azione di complessi informazionali integrati connessi alla coordinazione/inibizione.
- Modulazione della memoria: risultato dei processi che costituiscono i complessi informazionali integrati.
- Modulazione di soglia: modalità di costituzione del complesso informazionale integrato che modula il processo d’integrazione/elaborazione nel flusso di informazione nervosa. In questa costituzione si articolano i fenomeni bio-molecolari alla base del bilancio sensitizzazione/adattamento delle cellule nervose.
- Noxa: evento scaturito dall'interazione tra organismo e ambiente che induce perturbazioni su un bioritmo attraversandone un flusso informazionale. Nonostante l’etimologia del termine (che riporta ad un concetto di “danno” o di “lesione tessutale”) qui si riferisce unicamente alle dinamiche informazionali che tale evento produce indipendentemente dalle sue conseguenze patologiche immediate.
- Progetto di coordinazione: struttura del flusso informazionale che garantisce una corretta omeostasi tra dominio spaziale e temporale nella sua progressione sequenziale nel processo di integrazione/elaborazione. Consideriamo le manifestazioni patologiche come generali disequilibri di quest’istanza.
- Repertorio comportamentale: assetto predisposto di complessi informazionali integrati che modulano ai vari livelli di dominio spazio-temporale le leve funzionali rivolte all’esterno dell’organismo. Esso ha una componente variabile dovuta alla coesistenza del repertorio geneticamente determinato e del repertorio frutto di solchi mnesici individuali.
- Riserva simbolica: accumulazione di repertori di integrazione/elaborazione che esibiscono modalità simbolica/simbolizzazione nell’arco dello sviluppo evolutivo della specie e dell’individuo.
- Sensitizzazione/adattamento: fenomeno che consente a un sistema informazionale caratterizzato da alto dominio spazio-temporale (come il meccanismo di trasduzione analogico-digitale dei neuroni) la modulazione del rapporto stimolo-risposta alla base dell’accentuazione/riduzione del solco mnesico.
Gli argomenti della trattazione sono sintetizzati nei seguenti cinque punti.
1. Qualsiasi perturbazione (esogena o endogena) a carico dell’organismo produce una catena di eventi la cui entità, in termini di estensione anatomica e consumo energetico, è indipendente dalla grandezza dello stimolo. Tale discrepanza riflette l’azione computazionale dell’ospite che è in grado di incrementare o inibire le risposte allo stimolo, stereotipate e geneticamente determinate, lungo la loro stessa dimensione spaziale e temporale.
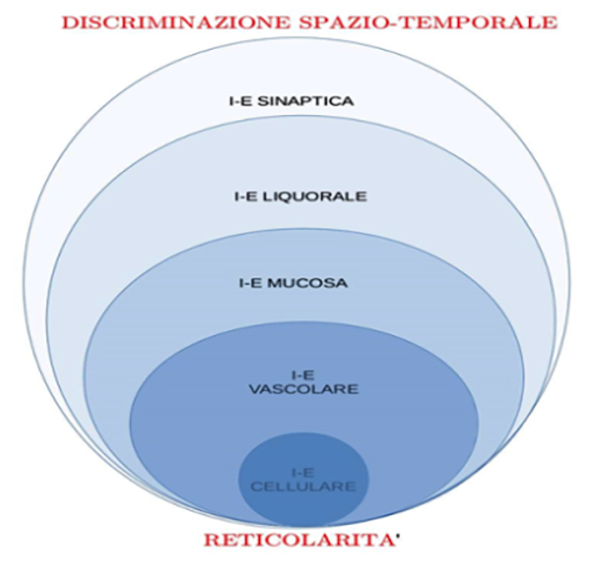
2. Le risposte alle perturbazioni ambientali avvengono ordinatamente lungo interfacce di scambio tra compartimenti adiacenti costituendo flussi di informazioni le cui cinetiche sono legate alla struttura tessutale e all’architettura tridimensionale di ciascun compartimento. I principali compartimenti, strutturati nell’arco della filo-ontogenesi, sono illustrati nella figura in basso ove “I-E” sta per “interfaccia intra-extracellulare”. All’apice troviamo la massima espressione di velocità di flusso: l’accoppiamento elettrochimico delle informazioni attraverso la codificazione analogico-digitale della sinapsi. Su tale dominio spazio-temporale, rapido e discriminativo, si verifica contemporaneamente la maggiore semplificazione del repertorio biochimico: poche molecole inserite entro interfacce protette dall’ambiente esterno “simboleggiano” la maggiore eterogeneità molecolare dei livelli di interfaccia sottostanti. Viceversa, alla base dell’ovoide si trova il massimo dell’eterogeneità biomolecolare: quella corrispondente alla prima interfaccia tra individuo e ambiente associata alla minima velocità di scambi e di discriminazione spazio-temporale (ovvero alla massima reticolarità di scambio di informazione). Il fatto che le diverse forme ovoidali siano immerse una nell’altra allude al fatto che tale modello interpretativo consente finalmente di raccogliere in un’unica disciplina organica la Neurologia, l’Endocrinologia e l’Immunologia.
3. Di fatto regolazione e computazione sono concetti coincidenti e va detto che i biosistemi computazionali sono dotati di memoria: influenzati da precedenti eventi perturbativi essi mutano l’entità della propria risposta in relazione alle caratteristiche temporali ed energetiche degli eventi stessi. La modulazione della memoria corrisponde all’attivazione di fenomeni biochimici peculiari che sovraintendono il bilancio tra sensitizzazione e adattamento. Il collegamento tra questo terzo assunto e il primo spiega da un lato le discrepanze tra entità dello stimolo ed entità della risposta e dall’altro la variabile latenza temporale tra perturbazioni e conseguenti manifestazioni fisiopatologiche (ad esempio: tachifilassi immunitaria o assuefazione per down-regulation postsinaptica).
4. Il succedersi di eventi perturbativi induce regolarmente entro il sistema fenomeni di riorganizzazione strutturale che si traducono in alterazioni irreversibili delle sue modalità computazionali.
5. Qualsiasi evento (generato da perturbazioni esterne o da dinamiche autopoietiche) è trasmesso a tutto il sistema. La sua manifestazione sul piano obiettivo deriva dal grado di eccitazione/inibizione scaturito da processi computazionali di ciascun livello di scambio tra compartimenti adiacenti. Salvo che per effetto di eventi particolari (ad es la folgorazione) il repentino blocco della trasmissione nervosa con Anestetico Locale (AL) costituisce un evento non prevedibile poiché assolutamente inusuale in natura. Il suo effetto corrisponde quindi alla massima istanza di riorganizzazione da parte del sistema biologico nella sua interezza e questo permette di spiegare la frequenza con cui l’applicazione terapeutica di AL, per quanto localizzata, può elicitare risposte globali.
Poco sopra si è fatto cenno al grado di chiarezza che vige intorno al background fisiopatologico delle Terapie Neurali e, consapevoli che molti elementi sono già patrimonio dei professanti, ci si concentra su quelli che ancora sembrano restare più nebulosi. Per introdurre il tema può risultare utile una metafora. Immaginiamo alcuni spettatori di venti minuti di football del tutto digiuni di conoscenze su quello sport. Dopo venti minuti essi avranno forse intuito che si fronteggiano due squadre distinguibili dal colore delle magliette e che il gioco, regolato da un tale in tenuta nera e munito di fischietto, consiste nel sospingere il pallone nella rete avversaria a forza di calci. Potrebbero anche intuire che il pallone non può essere toccato con le mani, che l’avversario non può essere percosso intenzionalmente e che al trillo del fischietto dell’uomo in nero il gioco si interrompe. Per quanto acuta sia stata l’attenzione degli osservatori non avranno acquisito che una minima parte delle competenze necessarie per comprendere pienamente il corpo complessivo di regole che governano il gioco poiché ancora privi di numerose conoscenze indispensabili. Tanto per cominciare: quali sono le dimensioni di un campo di football e quanto dura la partita? E’ evidente che aspetti cruciali delle grandezze dello spazio e del tempo non hanno potuto essere apprezzati. I giocatori corrono infatti per certe lunghezze in un certo tempo e inevitabilmente ciò li porta a esibire le rispettive capacità atletiche: scatto e velocità per alcuni, potenza o resistenza per altri. E poi, quali sono i ruoli dei singoli giocatori? C’è chi mantiene posizioni centrali ed effettua rapidi spostamenti per trasmettere il pallone con una logica variamente distributiva, chi resta in posizione arretrata e scatta quando un avversario si avvicina alla sua porta mentre altri ronzano nella metà campo avversaria risparmiando energie per farle esplodere al momento del passaggio del pallone sui loro piedi. Quanto pesa il pallone da football? Quanta potenza deve sprigionare un calcio per proiettarlo a trenta metri? Occorre convincersene: la comprensione del football non potrà che risultare parziale se ci si esime dal valutare le grandezze dello spazio e del tempo in cui il gioco si snoda e la forza, la velocità e la precisione necessarie nell’effettuare le evoluzioni previste da quello sport.
L’individuazione di numerosi organi all’interno dei viventi ha sempre suggerito l’idea di quest’ultimo quale contenitore di oggetti distinti e variamente collegati e diversi attori hanno via via impersonato tali collegamenti: dal modello cinquecentesco della circolazione ematica proposto da William Harvey agli ormoni circolanti, dal Sistema Nervoso al Sistema Immunitario. In epoca recente dalle corrispondenti discipline (Endocrinologia, Neurologia e Immunologia) sono emerse sempre più numerose connessioni tra entità biologiche che prima sembravano distanti sia funzionalmente che topograficamente. Progressivamente tali conoscenze hanno rivoluzionato la scienza medica dando vita alla Medicina Interna e aprendo un approccio parallelo al ragionamento medico e alla gestione del paziente. Infine l’impiego di moderne tecnologie e l’importanza di sensazionali scoperte in campo biomolecolare hanno potenziato lo studio dell’Embriologia illuminando il divenire della differenziazione e della collocazione anatomica tessutale nella costituzione dell’organismo maturo e rivelandone logiche architettoniche e analogie funzionali.
I primi dati sull’esistenza di comunicazioni tra i Sistemi Nervoso, Endocrino e Immunitario risalgono ad anni ormai lontani [42] e da allora le evidenze raccolte sempre più confermano l’esigenza di un nuovo modello medico sovraspecialistico. Hanno così tratto origine discipline “trasversali”: la Neuroendocrinologia, la Neuroimmunologia, la Neuropsicologia, la Psiconeuroendocrinoimmunologia. Offrendo sempre nuove conoscenze ognuno di questi passaggi ha incrementato il grado di complessità nel sapere biomedico ma ha anche proposto sempre nuovi nodi da sciogliere. Oggi conosciamo molti meccanismi grazie ai quali le emozioni condizionano le funzioni cardiovascolari e neuroendocrine e come queste stesse funzioni siano influenzate dall’esercizio fisico. Conosciamo molte modalità con cui elementi nervosi e immunologici interagiscono mediante scambi biochimici e informazionali in entrambi i sensi di marcia e sappiamo tanto altro ancora. Nonostante l’enorme incremento di conoscenze, specie quando una malattia si manifesta, si avverte l’insufficienza di coerenti applicazioni di un modello integrato: sembra quasi che la complessità dei singoli campi d’osservazione (biomolecolare, sperimentale, farmacologico, clinico) abbia prodotto nuove forme di segregazione culturale attraverso un’ulteriore moltiplicazione di ambiti scientifici tra loro difficilmente comunicanti.
Per giustificare quelli che talora appaiono come difetti di comprensione e/o di applicazione è lecito chiedersi quale sia il lato debole di questa nuova cultura medica trasversale. L’evidenza che linfociti, neuroni e ghiandole endocrine comunicano grazie a un repertorio condiviso di segnali non basta a soddisfare le nostre esigenze di sapere e saper fare. Recuperando il senso della metafora calcistica prevediamo che ulteriori passi conoscitivi e applicativi possano provenire dallo studio di quelle attività biologiche immerse nei dominii in cui si manifestano: spazio, tempo e intensità. La pratica quotidiana delle TAL “costringe” all’osservazione delle citate dimensionalità permettendo di cogliere alcune proprietà dei biosistemi regolatori che altrimenti resterebbero sfocate. Ma anche prescindendo dalla specificità della nostra attività professionale ogni altra figura che valuti un paziente deve rispondere a quesiti che chiamano in causa le citate grandezze se non altro poiché l’interpretazione è inevitabilmente dominata dal dualismo acuzie/cronicità. L’andamento acuto o cronico di una malattia (e anche l’alternanza di riacutizzazioni e remissioni nell’ambito di processi morbosi stabili) conducono a riflettere sull’estensione temporale dei fenomeni biologici di significato regolatorio. Da tempo la ricerca fisiopatologica esprime una massa enorme di dati la cui crescita è esponenziale. Per cogliere meglio il significato delle più rilevanti dinamiche biologiche oggetto di interesse se ne propone qui una lettura che preveda la loro collocazione entro il modello inclusivo offerto dalla combinazione dei dominii spazio-temporali: si suggerisce che la loro osservazione si declini nella valutazione del rapporto tra geometrie di flussi ed espressioni funzionali sollecitate da istanze regolatorie. Questo però richiede di deporre almeno parzialmente il criterio con cui solitamente vengono affrontati molti aspetti di Fisiopatologia. Sotto la luce che qui si propone può infatti risultare meno importante l’identità degli attori definita sulla loro origine embriologica (neuroni, immunociti, fibrociti, eccetera) rispetto alla loro collocazione entro i flussi, diversi per velocità e ampiezza, delle attività regolatorie. Per tornare alla nostra metafora calcistica: se un difensore percorre velocemente metà campo controllando il pallone fino a spingerlo in rete (gesto tipico di un attaccante) sarà la dinamica complessiva dell’azione nelle sue dimensioni spazio/temporali a meritare la nostra attenzione più che il numero esibito sulla maglietta del giocatore che ne indica il ruolo “ufficiale” nella squadra. Immaginiamo che l’adozione di questo modello sia utile a spiegare i principali fenomeni che caratterizzano la TNH (come il Campo di Disturbo = CdD e il Fenomeno del Secondo =FdS) e confidiamo nella bassa esposizione all’accusa di “eresia”. Esso infatti si colloca armoniosamente nella scia di un fronte di studi sempre più vasto inaugurato nel 1989 con la pubblicazione “A molecular basis for bidirectional communication between the immune and neuroendocrine systems” a firma dello statunitense Edwin Blalock [5]. Le evidenze che quello studio fece emergere imposero correzioni profonde nell’assegnazione dei ruoli: al Sistema Immune, fino ad allora identificato quale semplice aggressore di elementi non-self, vennero riconosciute prerogative di organo di senso e l’espressione di messaggi acquisibili dal Sistema Nervoso e a quest’ultimo, prima guardato come il campione della stabilità omeostatica, la capacità di suscitare processi morbosi esprimendo una messaggeria simil-immunologica.
Per definire alcuni elementi che useremo nella costruzione del nostro modello stabiliamo fin da ora cosa intendiamo per “dominio spazio-temporale”: esso è la combinazione tra la risoluzione spaziale e la risoluzione temporale in cui un evento si concretizza durante l’interazione con un osservatore. Per “risoluzione” si intende la distanza più piccola in cui possono essere distinti due punti adiacenti. Sul piano dello spazio il concetto è facilmente richiamato dai moderni televisori: a parità di superficie lo schermo che contiene più unità che emettono un segnale “pixel” presenta maggiore risoluzione spaziale rispetto a quello che ne emette meno: schermi di modesta superficie piana (11 pollici ad es) purché dotati di alta densità di pixel (ad es, 3840×2160 pixel) offrono immagini assai più nitide rispetto a schermi di 50 pollici (perciò ben più grandi) ma con una risoluzione di soli 800X600 pixel. Potrebbe risultare più arduo spiegare come lo stesso concetto si applichi alla dimensione temporale ma può tornare utile pensare alla tecnica con cui si producono i cartoni animati: facendo scorrere con la giusta rapidità disegni quasi identici ma dotati di lievi variazioni di posizione si ottiene una buona percezione di fluidità dei movimenti. Minori velocità di scansione delle variazioni delle posizioni o maggiori differenze tra la posizione di un dettaglio rispetto a quella del precedente potranno invece farci apparire il filmato meno fluido, procedente a “scatti”. All’estremo opposto la percezione della risoluzione temporale può essere apprezzata con l’impiego di tecniche di rilevazioni cinematografiche “lente” che, ad esempio, osservano la crescita e lo sviluppo di fiori e piante mediante la raccolta di milioni di fotogrammi per lunghi periodi di tempo. E’ in tale modo che i documentari scientifici ci fanno assistere alla crescita di germogli e allo sbocciare di fiori come se fossero vivacemente animati: in questo caso l’illusione deriva dal fatto di aver reso più “lento” lo strumento di raccolta delle scansioni temporali. Sono esempi questi che indicano come il tempo sia una dimensione percettiva, ossia relativa al rapporto tra oggetto e osservatore. Qui al centro dell’osservazione vi sono le modalità con cui le forme di vita hanno iniziato un percorso in cui prevalevano interazioni a bassa risoluzione spazio-temporale (sufficienti a scambi informazionali/nutrizionali nel brodo primordiale) dotandosi poi di modalità interattive sempre più “alte”. Queste ultime corrispondono a maggiori gradi di risoluzione spazio/temporale per competere in ambienti in cui la rapidità dei movimenti e la loro discriminazione nello spazio sono fattori cruciali per la competizione tra specie. L’assunto alla base di questa tesi è un elemento pienamente accolto dalla Comunità Scientifica che tuttavia spesso viene trascurato: esso prevede che le strutture ancestrali deputate all’interazione su bassi livelli di risoluzione spazio/temporali non siano mai state scartate nel corso dell’evoluzione ma “conservate” in una condizione di crescente controllo da istanze inibitorie progressivamente sovrastanti. Nella nostra ricostruzione cercheremo di individuare strati sovrapposti di tali domini spazio-temporali nell’architettura geometrica e funzionale degli organismi viventi. Descrivere con questa chiave di lettura aspetti rilevanti di Fisiopatologia rende opportuno l’impiego di alcuni termini inediti ed è per questo che prima è stato offerto un breve glossario.
30a.
Un nostro parente prossimo: l’Hydra Linnaeus
Già dai primi decenni del XX° Secolo iniziarono ricerche in vivo su modelli sperimentali rappresentati da rettili, anfibi e animali della zoologia superiore ma quando si vogliano comprendere aspetti fisiopatologici emersi dai gradini più profondi dell’evoluzione, come nel nostro caso, può risultare utile partire dall’osservazione di forme di vita ancora più semplici e antiche. Questi studi fanno emergere alcuni aspetti della Filogenesi e dell’Embriologia che consentono un approccio organico alla materia fisiopatologica nel suo insieme e al tempo stesso conducono alla comprensione delle basi delle forme di neuro-immunomodulazione perseguite con le Terapie con Anestetici Locali (TAL).
L’Hydra, uno dei più semplici tra i sistemi pluricellulari, presenta un sistema contrattile che permette la diffusione di uno stimolo attraverso la contiguità di più cellule secondo una modalità “sinciziale”: stimoli a partenza dalla sua periferia somatica attraversano diffusamente e simultaneamente tutto il suo organismo fino a promuovere un comportamento globale di apertura/chiusura. In un simile sistema le sedi di partenza e di arrivo dell’onda informazionale-contrattile tendono a coincidere e l’area somatica preposta all’esplorazione ambientale è in genere la stessa che attua l’ingestione (la corona di propaggini che funge da bocca). Il destino del materiale fagocitato è legato alla sua composizione biochimica. Rispetto al rapporto tra l’esigenza di sopravvivenza e l’offerta biochimica entra così subito in gioco la capacità di discernimento. Durante la filogenesi delle specie viventi il discernimento costituisce l’istanza evolutiva principale dei processi di differenziazione cellulare: per approfittare della disponibilità di un maggior numero di alimenti gli organismi pluricellulari hanno espresso altrettanti nuovi compartimenti per l’analisi/fruizione delle loro caratteristiche biochimiche per veicolare sostanze semplificate a tipi cellulari diversificati. Emerge qui un forte cuneo evolutivo: effettuare un maggior numero di elaborazioni biochimiche del materiale alimentare induce la necessità di nuovi e diversi tipi cellulari ma ciò richiede un incremento del volume corporeo con esposizione a più rischi di attacco dall’esterno o di collasso energetico (un numero troppo alto di cellule a parità di offerta nutrizionale).
Nel corso dell’evoluzione la strategia adattativa di molte forme di vita primordiali, tra cui l’Hydra, ha assunto quattro modalità plastico-funzionali: l’elongazione, l’avvoltolamento, la cefalizzazione (procedendo dal basso verso l’alto quest’ultima integra le prime due fasi con la spinta di architetture pluricellulari sempre più avvoltolate lungo l’asse longitudinale) e infine l’assunzione di competenze locomotorie [8]. Tali modalità evolutive si sono variamente intrecciate nelle diverse specie ma, globalmente, si sono espresse con una spinta locomotoria coincidente con l’orientamento della cefalizzazione. Per comprendere i passi successivi della trattazione sarà necessario identificare tali quattro modalità plastico-funzionali, quale “patrimonio conservato”, negli animali della zoologia superiore e nell’uomo. Dall’osservazione di questi processi già emergono quattro rilevanti corollari ma prima di enunciarli esortiamo l’Utente, ove questo elemento difetti, al pieno recupero del significato di cosa significhi “conservazione” nelle dinamiche evoluzionistiche. A questo fine suggeriamo la lettura del capitolo (32) e la visione del filmato (FEF3).
Il primo corollario vede la coincidenza della nutrizione con la perlustrazione dell’ambiente. La competizione tra individui implica però anche la produzione di sostanze tossiche indovate nei nutrienti fagocitati e allora potrebbe essere non per caso che la gran parte delle malattie infettive non virali dell’uomo sia sostenuta da tossine prodotte da microorganismi entrati per via gastroenterica o inalatoria. In ambo i casi gli agenti infettanti, mossi dalla necessità di colonizzare l’ospite, sfruttano l’interfaccia mucosa di procacciamento di nutrienti. E’ da questo ambito della vita di relazione tra specie che trae origine l’ancestrale interrelazione tra Sistema Nervoso (SN) e Sistema Immunitario (SI).
Il secondo corollario prevede che il flusso di informazioni corrispondente alla nutrizione avvenga da un lato sul piano “contrattile” (il dispositivo neuro-muscolare che coordina la fagocitosi), dall’altro lato sul piano “biochimico” (il dispiegamento nell’organismo di sistemi di analisi/fruizione degli alimenti). Durante la filogenesi e l’ontogenesi i due flussi informazionali, pur viaggiando in diverse risoluzioni spazio-temporali, rimangono costantemente paralleli. Questo aspetto è cruciale per comprendere rilevanti implicazioni fisiopatologiche e cliniche e già possono scorgersi distinti ambiti architettonici in cui i due flussi di informazione trovano spazio: il flusso d’informazione contrattile si disporrà in un sistema sinciziale di cellule contigue a formare fasci di connessione tra punti diversi dell’organismo ove la contrazione in un punto indurrà la contrazione delle cellule contigue attraverso un accoppiamento elettro-chimico tra citomembrane adiacenti (si tratta del “prototipo” del SN). Il flusso di informazione biochimico è rappresentato dalle sostanze disciolte nel mezzo fluido extracellulare e risulta costituito dalla matrice glicoproteica idrofila che definisce un ambiente reticolo-lamellare più o meno denso (“stroma”). Man mano che si formano compartimenti pluricellulari più complessi si creano cavità comunicanti sempre più liquide e a scorrimento più veloce delimitate da cellule più specializzate dette “endoteliali”. Il passaggio è confermato dall’origine mesenchimale della linea emato-linfopoietica in una prima fase dai connettivi reticolari dei visceri annessi all’intestino (fegato e milza) e poi da quelli delle ossa lamellari (midollare ossea). Entro il flusso d’informazione biochimica si riscontrano tutte le istanze nutrizionali e, sulla scorta di quanto premesso, al tempo stesso si realizza il primo abbozzo di competenza immunitaria.
Con il terzo corollario si può affermare che le relazioni tra i flussi informazionali non siano paralleli ma complanari: a differenza delle linee parallele della geometria euclidea tra tali flussi complanari esistono punti d’incontro. Un processo di integrazione-elaborazione avviene immediatamente, diffusamente e ubiquitariamente dove l’interfaccia è fornita da uno strato di cellule contrattili (i miociti, cellule starter del “flusso contrattile”) e dall’ambiente stromale (“flusso biochimico”) con l’intermediazione di uno strato di cellule endoteliali. Queste ultime sono immerse nello stroma con la funzione cruciale di “interprete”. Allo stato di contrazione dei miociti corrisponde un dato consumo energetico registrato dalle cellule endoteliali come flusso di informazione biochimica. Le cellule endoteliali rilevano il tenore energetico dell’interfaccia miocita/fluido (in termini di consumo di ossigeno e produzione di cataboliti) producendo un catabolita che si materializza e si distrugge nell’arco di millisecondi e che induce l’immediata riduzione dell’attività contrattile dei miociti: l’ossido nitrico. La decontrazione che esso induce rilassa lo stroma in cui sono immerse le cellule endoteliali producendo il loro reciproco allontanamento e questo incrementa il flusso dei nutrienti tra le cellule endoteliali anzitutto a beneficio energetico dei miociti. Tale processo spiccatamente autorigenerativo (contrazione / consumo energetico / ossido nitrico / decontrazione / incremento del flusso energetico / contrazione) è costante, ripetitivo e primario: viene cioè innescato e perpetuato dalla presenza stessa degli attori citati mostrando le caratteristiche di “bioritmo autogeno”. Questi fenomeni basali rientrano nella definizione generale di “vasomotion” dal cui studio è possibile distinguere le attività oscillatorie influenzate da fattori nervosi, ormonali, immunologici e locali. Quella appena descritta è la “vasomotion related to endothelial activation” [53], dispositivo biomolecolare di autoregolazione che attraversa longitudinalmente la linea filogenetica da forme di vita molto semplici fino all’uomo (che ha trovato il modo di correggerne alcune disfunzioni impiegando i nitrati nell’angina pectoris e il Sildenafil nella disfunzione erettile).
Nel flusso informazionale biochimico non è contenuto solo l’apporto energetico dei nutrienti disponibili localmente (glucosio, lipidi, aminoacidi) ma anche informazioni relative all’ambiente esterno e/o allo stato energetico di distretti somatici più lontani. Perciò localmente le cellule endoteliali acquisiscono competenza anche rispetto a sorgenti remote d’informazione e prendono a produrre altre sostanze capaci di decontrarre i miociti. Esempi di tali modalità di amplificazione funzionale endoteliale sono la Bradichinina, la Prostaciclina e il Fattore di Depolarizzazione Derivato dall’Endotelio (EDHF). La relazione tra i due flussi di informazione (quello contrattile protonervoso e quello biochimico costituito dalle sostanze disciolte nello spazio intercellulare) sostiene quindi la funzione di integrazione-elaborazione. Intendiamo quest’ultima quale capacità di amplificare o ridurre funzioni di un sistema già autoregolato dietro istanze anche assai distanti da un dato settore somatico. Ad esempio la secrezione di Bradichinina nel torrente siero-ematico (da qualunque causa sia indotta) produrrà sul piano informazionale una perturbazione globale del bioritmo autogeno mentre sul piano funzionale determinerà l’incremento del flusso siero-ematico locale.
Tornando ai rapporti tra morfologia e funzione il menzionato avvoltolamento assume un’importanza cruciale nella complessità dei sistemi viventi pluricellulari. Il processo filo-ontogenetico che lo sostiene è l’elongazione, espressione dell’incremento di processi informazionali lungo un percorso longitudinale (ovvero il moltiplicarsi di processi biologici sequenziali lungo una linea). Tanto più lungo sarà il percorso quanto maggiore si esprimerà il livello di complessità del flusso informazionale del corrispondente dominio spazio-temporale. In altre parole: il flusso nutrizionale nel canale alimentare è tanto più ricco di eventi perturbativi-regolatori quanto maggiore è la sua lunghezza. Se gli organismi viventi fossero rimasti confinati unicamente nella dimensione della lunghezza (esprimendo al contempo la variabilità e la complessità delle operazioni molecolari e rispettandone la sequenzialità temporale) oggi ci apparirebbero mostruosi: un neurone cerebrale sarebbe dislocato a diversi chilometri dal suo effettore finale, ad esempio un muscolo del piede, e le dimensioni dei nuclei cellulari (che ospitano diversi metri di materiale genomico) risulterebbero enormi per l’incapacità di avvoltolarsi nella costituzione elicoidale dei cromosomi. In ossequio a questo universale criterio di economicità le sedi dei processi di integrazione-elaborazione assumono forma tanto più avvoltolata quanto più ampio è lo scambio tra flussi informazionali. L’intestino costituisce un esempio lampante di avvoltolamento ed è facile intuire quanto questa disposizione spaziale favorisca il governo neuroendocrino delle sue numerose funzioni rispetto a come verrebbe esercitato se invece fosse disposto in forma elongata.
Proprio in questo contesto emerge con forza il ruolo informazionale di un livello di struttura-funzione a cui finora non abbiamo fatto cenno. Il dominio spazio-temporale dell’interfaccia sierosa rappresenta esattamente il substrato strutturale dell’avvoltolamento di vie longitudinali. La pleura, il peritoneo, la capsula glissoniana epatica, il pericardio e, soprattutto, il rivestimento meningeo del Sistema Nervoso Centrale (SNC) delimitano lo spazio di contenimento del processo di avvoltolamento dei rispettivi piani di dominio sottostanti (le vie di flusso nutrizionali, contrattili e nervose) e, al tempo stesso, si costituiscono quali unità funzionali compatte dotate di autonomia di risposta e “rappresentative” del loro contenuto viscerale. E’ proprio per questo che, ad esempio, la sollecitazione di un punto del rivestimento peritoneale induce una perturbazione contrattile e secretiva tendenzialmente estesa a tutto il tratto intestinale (si pensi al blocco della peristalsi in corso di appendicite) così come la presenza di un focolaio infiammatorio aracnoideo coinvolge tutte le funzioni nervose centrali pur in assenza di una sofferenza diretta dei neuroni del nevrasse. L’avvoltolamento del contenuto di un dominio spazio-temporale entro il suo contenente sieroso implica il compattamento di una via longitudinale entro uno spazio tridimensionale e ciò induce necessariamente la formazione di siti trasversalmente adiacenti in cui i punti di contatto sono separati dalle strutture parietali della via informazionale stessa. Tanto più tale disposizione si presta al passaggio trasversale di perturbazioni (nutrienti, citochine, ormoni, sequenze di potenziali d’azione) attraverso i punti di contatto tra anse adiacenti della via di flusso, quanto più la direzionalità delle informazioni tende a perdere la continuità della sua sequenzialità longitudinale. La stessa costituzione geometrica tridimensionale delle architetture biologiche offre un sistema regolatorio generale capace di modulare il bilancio tra flusso informazionale longitudinale e un contemporaneo flusso trasversale in base a istanze adattative: ne è un esempio la regolazione del bilancio idro-elettrolitico tramite il percorso del filtrato glomerulare lungo il gradiente acido-basico corticomidollare che l’ansa di Henle compie nel tubulo renale. Si comprende facilmente quanto possa risultare disastrosa l’evenienza di un sovvertimento patologico di questo bilancio di flussi tra loro trasversali. Malattie drammatiche e irreversibili, come la cirrosi epatica, si caratterizzano proprio dalla perdita della demarcazione tra flussi adiacenti nell’ambito di una stessa unità funzionale.
Allorché la sovrapposizione di effetti derivanti da diverse istanze ravvicinate induca la sollecitazione dello stesso bioritmo si rende necessaria la presenza di segregazioni fisiche che inibiscano il passaggio di un numero troppo elevato di flussi informazionali su un singolo sito di integrazione. Quanto maggiore è il livello di avvoltolamento (ossia la segregazione in uno spazio tridimensionale “piccolo” in rapporto all’incremento della lunghezza della via informazionale) tanto più difficilmente valicabili dovranno essere i confini rispetto all’eventualità patologica di un travaso trasversale tra anse adiacenti. Il sistema che mostra la maggiore espressione ontogenetica di avvoltolamento si riscontra nella struttura istologica della corteccia cerebrale: ogni circonvoluzione è separata dall’altra da tre rivestimenti: lo strato delle cellule endoteliali dei capillari (tra loro adese tramite thight juction, la cosiddetta “barriera ematoencefalica”), il liquido intercellulare dello spazio di Virchow-Robin e, infine, lo strato basale delle cellule astrocitarie sottostante alla pia madre. All’interno di questa stratificazione si articolano tutte le relazioni funzionali tra ambiti extra- e intranevrassiali per il concorso di innumerevoli relazioni tra regolazione dei flussi ematolinfatici, di quelli liquorali e i passaggi di citochine infiammatorie. Le circonvoluzioni cerebrali forniscono la prova di come l’istanza filo-ontogenetica dell’avvoltolamento abbia raggiunto l’estremo gradino dell’evoluzione e di come la sua incompleta realizzazione inevitabilmente si traduca in disfunzioni sul piano clinico: a disordini della migrazione neuronale (come la pachigiria e la lissencefalia) in cui sono evidenti difetti di vario grado di avvoltolamento, si associano regolarmente epilessia, ritardo mentale e altri disordini [4]. La trattazione della fisiopatologia di questo complesso distretto somatico apre a numerosi spunti di riflessione sulle relazioni intercorrenti tra informazione, infiammazione e strutture geometriche, temi questi che affronteremo impiegando modelli più semplici e che, a conclusione di questo scritto, ritroveremo come base morfologica e funzionale di una delle più sconcertanti scoperte della Neurofisiopatologia degli ultimi anni: il “sistema gliolinfatico” e le sue implicazioni in alcune delle patologie del Sistema Nervoso Centrale più temute nella nostra epoca: il Morbo di Alzheimer e la Sclerosi Multipla.
Siamo al quarto corollario. La convergenza nei punti di integrazione-elaborazione può essere modulata da istanze provenienti da entrambe le sorgenti informazionali: perturbazioni del sistema contrattile influiscono sui processi di integrazione-elaborazione analogamente alle perturbazioni biochimiche. E’ noto ad esempio che le discinesie gastroesofagee, sostenute da fenomeni pertinenti alle dinamiche disfunzionali della digestione, possano indurre turbe del ritmo cardiaco per via della comune appartenenza allo stesso plesso nervoso vegetativo. La differenza di risultato tra una perturbazione di origine contrattile e una di origine biochimica si riverbera però su due dominii: il tempo (più rapido o più lento) e l’estensione spaziale (puntiforme o diffusa), cioè esattamente sulle due dimensioni individuate come caratteristiche proprie dei flussi informazionali. Se ne deve concludere che la sostanza che diviene attrice del processo di integrazione-elaborazione (nel nostro esempio la Bradichinina) non è soggetto peculiare di una specifica finalità fisiologica ma rappresenta la “leva” di un meccanismo che può essere innescato da fonti informazionali a velocità ed estensioni diverse e su istanze nutrizionali-comportamentali potenzialmente anche divergenti. Quanto a tipologia molecolare le varie “leve funzionali” identificate nell’uomo (neurotrasmettitori, neuromodulatori, citochine) risultano ubiquitarie in regioni dell’organismo remote e funzionalmente lontane: la stessa molecola di serotonina è presente nei granuli citoplasmatici delle piastrine circolanti come nelle sinapsi dei nuclei del rafe del Ponte di Varolio. Più che frutto di scarsa fantasia genomica tale economia biosintetica comprova un processo di migrazione di strutture architettoniche ancestrali in prossimità topografica e funzionale con strutture evolutivamente più recenti.
Concludiamo il paragrafo rimarcando un primo concetto utile a formulare un aspetto della Fisiopatologia dei biosistemi di regolazione: indipendentemente dalla composizione biochimica, dalla sede anatomica e dai meccanismi molecolari che può innescare definiamo “leva funzionale” qualsiasi attore di interazione integrativo/elaborativa tra flussi informazionali complanari.
30b.
La comparsa nei viventi della “coordinazione-inibizione” attraverso la differenziazione delle vie informazionali.
È arduo individuare tutte le logiche alla base dell’evoluzione dei sistemi biologici perché l’analisi delle forme di strutturazione delle specie pluricellulari dovrebbe tenere in rassegna le innumerevoli manifestazioni della biodiversità partendo da un inizio della vita di cui non si conosce alcuna data certa. Negli animali della zoologia superiore ritroviamo però le caratteristiche architettonico-strutturali dell’Hydra nel dispiegamento spaziale del canale alimentare: elongazione, avvoltolamento e cefalizzazione (quest’ultima corrispondente all’integrazione tra livelli di elaborazione ordinati gerarchicamente). L’estremità cefalica e quella caudale costituiscono i punti di convergenza tra un sistema contrattile automatico e un sistema intenzionale identificandovi i due poli di integrazione/elaborazione neuro-muscolare. Non sembra perciò casuale che la loro innervazione spinale sia affidata a nuclei nervosi (bulbare il primo e sacrale il secondo) in forme di vita più evolute definiti “parasimpatico cranio-sacrale” quasi a sottolineare che in sostanza si tratta di un tutt’uno. Per afferrare il senso di questa disposizione bisogna richiamarsi ai movimenti dei flussi di informazione che avvengono sincronicamente e su assi geometricamente interconnessi. Il flusso d’informazione che guida tutti gli altri è quello che assicura la sopravvivenza dell’individuo: l’apporto nutritivo. Trascurando il sistema integrativo che governa la scelta di cosa ingerire, l’atto dell’ingestione e il semi-automatismo della deglutizione la via informazionale biochimica assume il ruolo di iniziatore e moltiplicatore di processi informazionali complanari su punti di integrazione/elaborazione sequenziali (cioè inseriti lungo il tragitto del cibo). Infatti ciò avviene dal tratto esofago-gastrico fino alla valvola ileo-cecale ove il ruolo di interprete è affidato ai recettori chemio-fisici presenti nell’epitelio. Ogni tipo molecolare assunto (glicidi, aminoacidi, lipidi, ioni, eccetera) attiva separatamente vie informazionali complanari distribuite ciascuna in un proprio dominio spazio-temporale, cioè in tempi e su estensioni diverse in ambito paracrino (diffusione entro lo stroma e le vie linfatiche), endocrino (diffusione entro vie ematiche) e nervoso (diffusione entro vie di conduzione elettrochimica in sede mioenterica, gangliare, spinale ed encefalica). Una fase digestiva corrisponde perciò all’attivazione dell’intero organismo secondo una sequenza longitudinale (corrispondente al tragitto del nutriente) e una sequenza trasversale (corrispondente all’estensione delle vie informazionali reclutate che si spingono a macchia d’olio dall’interfaccia mucosa all’encefalo). L’immagine va integrata con la spinta filo-ontogenetica dell’avvoltolamento del tubo intestinale ove, mentre ciascuna via informazionale persegue le proprie specificità funzionali nella rispettiva estensione spazio-temporale, si verifica la sovrapposizione di più estensioni. Il tutto avviene lungo un periodo corrispondente a un ciclo di passaggio del nutriente lungo la via informazionale biochimica iniziale, quella del cibo.
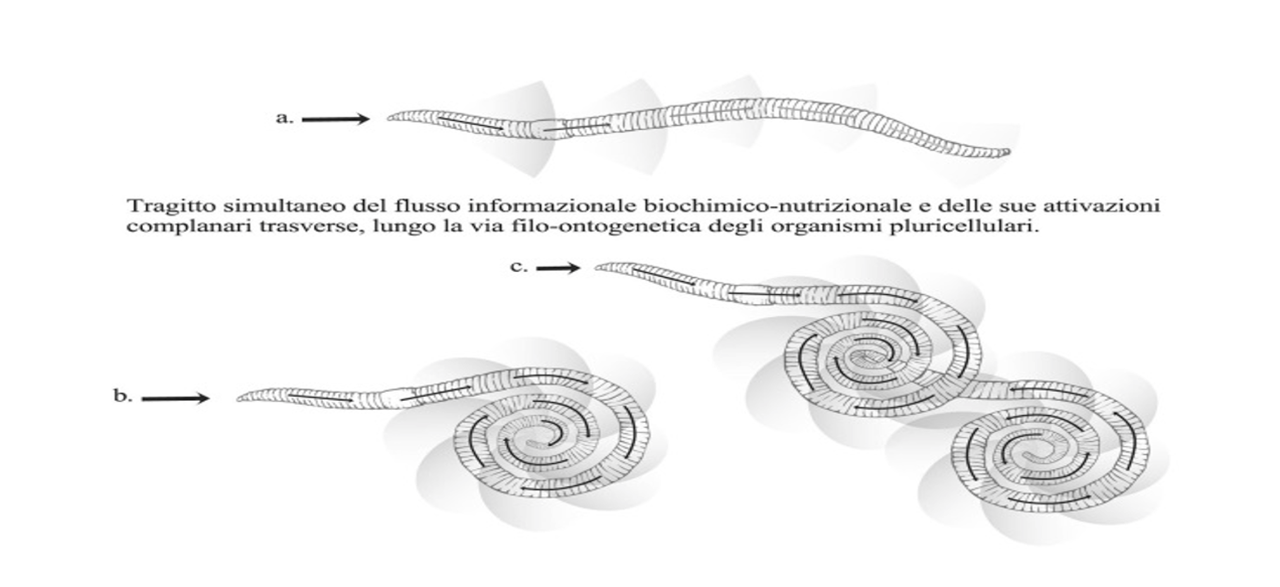
Per inserire correttamente in questo contesto l’emergenza della funzione immunitaria va considerato che il cibo possa costituirsi anche come “nemico esterno”. A tal proposito è suggestivo che una vasta letteratura sulla filogenesi delle lipoproteine plasmatiche indichi un loro ruolo ancestrale quali mediatrici dell’infiammazione, ruolo che a sua volta si connette al loro transito entero-epatico [30]. E’ noto che la degradazione intestinale delle membrane batteriche costituisce l’atto preliminare all’assorbimento dei lipidi, la fonte energetica più ricca in natura. L’istanza di sopravvivenza non permette di scartare alcuna fonte nutritiva per cui anzitutto si assumono tutte le risorse possibili e poi, semmai, ci si difende dalle eventuali insidie in esse indovate. Ciò permette di postulare una sorta di identificazione tra il flusso di informazione biochimico nutrizionale e il suo contenuto tossico-infettivo. Ora occorre individuare le sedi di integrazione/elaborazione in cui il flusso tossico-infettivo giunge a contatto con le vie informazionali complanari superiori. In ambito paracrino troviamo il tessuto fibroreticolare dello stroma epatico (cellule di Kupfer) e delle linfoghiandole intestinali. In ambito endocrino (nel flusso ematico) troviamo le cellule granulocitarie. Nell’ambito che abbiamo definito “pre-nervoso” troviamo da una parte i linfociti che si attivano mutandosi in cellule contrattili e secernenti immunoglobuline (plasmacellule) e dall’altra linfociti che talora fungono da recettori interpreti mentre altre volte da produttori di leve funzionali (citochine e ormoni). Poiché scaturita da una via d’accensione longitudinale (immaginiamo il percorso del cibo simile alla combustione di una miccia) l’attivazione trasversale conduce alla sovrapposizione di più vie informazionali che attivano allo stesso tempo più leve funzionali. Una via informazionale “a bassa risoluzione spazio-temporale” sarà topograficamente estesa, lenta nella sua propagazione e quindi anche nell’attivazione delle sue leve funzionali e altrettanto lento sarà l’esaurimento dell’effetto funzionale di queste ultime.
E’ il momento di introdurre il concetto di inibizione connessa alla coordinazione. Mentre non è possibile identificare settori funzionalmente distinti in organismi primordiali come l’Hydra, nella zoologia superiore il sistema enterico origina dalla differenziazione di aree dell’organismo dedite contemporaneamente a più attività. Ma l’attivazione contemporanea di più vie informazionali può confliggere con l’efficienza generale del sistema: partecipare a una maratona dopo un pranzo sontuoso è impresa ardua così come compiere un profondo atto respiratorio contemporaneo a un atto di deglutizione. Per “coordinazione” si deve intendere che una stessa produzione di attività, col suo contenuto informazionale ed energetico, si disloca allo stesso tempo secondo una sequenzialità temporale e una discriminazione spaziale. Per una migliore comprensione impieghiamo una metafora cinematografica. Per trasformare più fotogrammi in una sequenza cinematografica dobbiamo garantire per ognuno di essi lo stesso grado di risoluzione (tanti punti in tanto spazio) pur modificando nel tempo la distribuzione dei punti che delineano il cambiamento delle figure. I processi elettrochimici insiti nei circuiti nervosi sono strutturati in modo da esibire questa capacità. Uno schema semplice è quello dei circuiti di relè interposti nelle stazioni intermedie delle vie lunghe spinali: più terminazioni assonali convergono su una fila di neuroni in un dato rapporto numerico (tot assoni proiettano su tot neuroni) tale per cui il numero di neuroni che raccolgono i potenziali trasmessi dagli assoni può variare in senso convergente (numerosi assoni e pochi neuroni) o divergente (pochi assoni e numerosi neuroni). Alla stessa disposizione architettonica del circuito corrisponde immediatamente l’elaborazione informazionale (espansione o contrazione dell’immagine trasmessa). Premesso che regolazione e computazione sono concetti coincidenti le singole connessioni tra assoni e neuroni sono di fatto configurate nella forma più idonea a esprimere la capacità di effettuare attività algebriche: un assone conduce “informazione +1”, l’altro conduce “informazione -1” e quindi il risultato è “informazione 0”. Alla possibilità di espansione-contrazione dell’immagine informazionale si aggiunge così quella del contrasto: oltre che ingrandire o ridurre un’immagine si vengono a stagliare al suo interno delle forme. L’articolazione nel tempo di diverse operazioni aritmetiche entro circuiti nervosi è un fenomeno squisitamente centrale e, infatti, le sinapsi inibitorie che permettono la sottrazione algebrica compaiono come caratteristica del “nevrasse” che rappresenta il settore di SN filogeneticamente più moderno.
Siamo al quinto corollario. Perché funzioni di coordinazione possano svolgersi correttamente i competenti centri di elaborazione devono disporre di tutte le informazioni necessarie a riformulare un progetto di coordinazione. Tornando alla metafora cinematografica: non è possibile assemblare un filmato partendo da fotogrammi con diverse risoluzioni. I sistemi di flusso informazionale elettrochimico saranno allora implicati nel trasporto trasversale di tutte le informazioni concernenti l’ambiente locale di partenza fino a un luogo di integrazione/elaborazione situato su un piano di flusso informazionale nervoso più elevato. Stiamo alludendo alla “centralizzazione funzionale”, processo fondamentale della fisiologia nervosa. Per soddisfare questa esigenza nel suo sviluppo verso l’alto il Sistema Nervoso prende con sé settori di periferia innervata per rappresentarli “in modalità simbolica” entro la sua capacità computazionale. A base della moderna Neurofisiologia questo è rappresentato nell’immagine dell’omunculus della corteccia pre- e post-rolandica dell’Homo Sapiens. I diversi settori di periferia innervata, tradotti in segnali nervosi digitalizzati, si costituiscono in flussi complanari di informazione. Per un verso essi risultano liberati del “peso analogico” della loro specifica natura chimico-fisica mentre, per altro verso, sono “costretti” a trascinare in direzione cefalica le informazioni di tutte le vie biochimiche e biofisiche scaturite dalle loro interazioni con l’ambiente. Tali vie si sono sviluppate su diverse risoluzioni spazio-temporali e, per questo, le gerarchie nervose sono rappresentative di tutte le diverse risoluzioni spazio-temporali attraverso la trasmissione complanare di vie afferenti ed efferenti a diversa velocità di conduzione, a diversi gradi di interconnessione sinaptica e con diversi campi recettoriali-effettoriali.
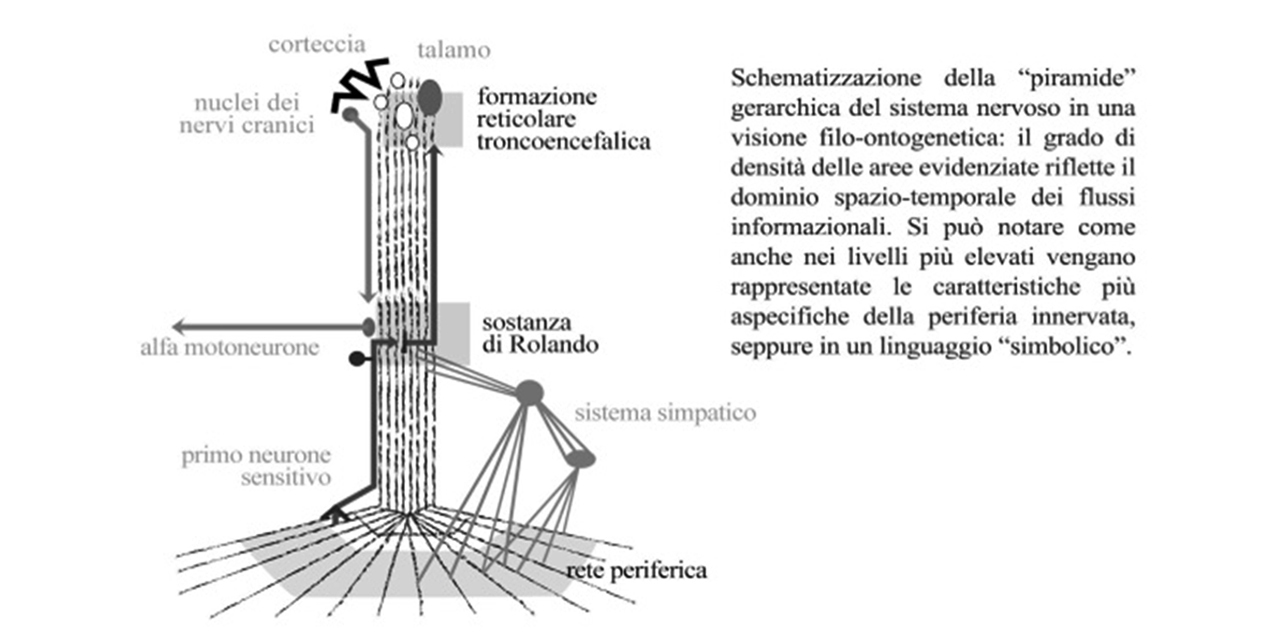
Questa architettura corrisponde a una sovrapposizione di risoluzioni spazio-temporali sempre più alte in una piramide il cui apice è largamente permeato dai livelli sottostanti. La capacità di esprimere funzioni d’insieme è garantita dai processi di coordinazione/inibizione scanditi sul piano temporale. Fenomenologicamente ciò corrisponde alla sovrapposizione dei bioritmi nella costituzione di quelle attività che convenzionalmente (ma impropriamente) chiamiamo “toniche”. Nel ritmo sonno-veglia troviamo una splendida rappresentazione di attivazioni lente scandite da sostanze che seguono una pulsatilità di ore (come la melatonina e l’oressina) sovrastate da ritmologie rapide dei neuroni del sistema reticolare ascendente che scandiscono l’alternarsi di accensioni-spegnimenti dell’ordine di minuti. Le interrelazioni tra questi dominii (“ormonale” il primo e “nervoso” il secondo) sono mediate da fenomeni inibitori che ne contrastano il possibile conflitto reciproco. Si struttura così un’architettura temporale in cui tutti gli attori, sia quelli rapidi che quelli lenti, sono costantemente ma variabilmente coesistenti.
3c.
La “centralizzazione” del Sistema Nervoso trascina in alto anche competenze immunitarie?
E’ stato premesso che la fagocitosi costituisce un punto di incontro tra funzione nutritiva e Sistema Immunitario (SI) e che i processi di sviluppo evolutivo coinvolgono entrambe le finalità funzionali. Si apre allora un quesito: se il SN ha trasferito verso l’alto settori della periferia somatica tradotti in simboli possiamo immaginare che esistano altrettanti livelli gerarchici del SI correlati a quelli nervosi? Nella struttura biomolecolare dei gangli delle catene ortosimpatiche pre- e laterovertebrali troviamo rappresentate in linguaggio nervoso attività di riconoscimento, memorizzazione e di leva funzionale (produzione di sostanze vasoattive e citochine infiammatorie) rispetto al flusso di eventi biochimici che pervengono al ganglio competente su quel campo recettoriale [9][16]. Inoltre la superficie dei linfociti, i più evoluti effettori della risposta immunitaria, esibisce recettori adrenergici, purinergici, glutammaergici e istaminergici: dispone praticamente di tutta la recettorialità necessaria alle funzioni di integrazione/elaborazione tra i flussi informazionali prima descritti [19][25][27][33][35][52].
Il processo di coordinazione-inibizione si articola nella struttura d’insieme sia sul flusso d’informazione nervosa (con l’interposizione di meccanismi inibitori) sia su quello biochimico (con i fenomeni di tolleranza immunitaria). E’ stato anche detto che il punto d’incontro tra flussi informazionali da cui scaturisce l’elaborazione-integrazione produce una perturbazione del bioritmo globale. Ma se, alla luce della iniziale definizione di dominio spazio-temporale, ci limitassimo alla misurazione temporale del bioritmo trascurandone la dimensione spaziale rischieremmo di non comprendere le relazioni alla base dei fenomeni fisiopatologici per difetto di globalità dell’osservazione. Spesso le manifestazioni patologiche fungono da evidenziatori della realtà mentre quelle fisiologiche più facilmente mostrano il “mare calmo” degli equilibri omeostatici. Il progressivo perfezionamento delle capacità informazionali del SI emergono da studi sull’evoluzione che hanno individuato la variabilità dell’espressione fenotipica delle immunoglobuline e dei recettori linfocitari quali espressioni di progressivi adattamenti sviluppatisi nella filogenesi e nell’ontogenesi [11][51]. Emerge una forte simmetria di sviluppo tra i due sistemi: in quello nervoso si realizza una segregazione di più vie complanari tra vie discriminative oligosinaptiche ad alta velocità e vie reticolari multisinaptiche a bassa velocità di conduzione mentre quello immunitario si porta contemporaneamente su diversi vettori entro vie, più o meno liquide, dallo stroma connettivale al sangue passando per le vie linfatiche. Analogamente, i processi maturativi e differenziativi che seguono all’interazione antigene-anticorpo avvengono entro strutture reticololamellari (linfonodi, milza, fegato e midollo osseo) in cui le linee maturative delle cellule immunocompetenti si sviluppano lungo linee parallele e segregate. All’incremento della segregazione architettonica dei punti di integrazione/elaborazione corrisponde l’incremento della quota dei processi inibitori elettrochimici. In altri termini: tanto più il raggruppamento dei domini spazio-temporali diviene compatto lungo la linea ascendente della cefalizzazione quanto più esso risulta dotato di dispositivi coordinativi-inibitori. Tale modalità architettonico-funzionale è descritta magistralmente nella monografia di sir John Carew Eccles sulla fisiologia del cervelletto, struttura encefalica in cui i fenomeni di coordinazione-inibizione articolati nella rappresentazione centrale del movimento emergono con particolare chiarezza [15]. Nello sviluppo filo-ontogenetico la combinazione finale dei due processi maturativi giungerà a un determinato grado di coordinazione dell’attività globale del biosistema, dato questo che risulta evidente nella disposizione per metameri di competenze attinenti a diversi settori di periferia somatica. Entro ciascun metamero il complesso di flussi informazionali viene segmentato nella citata architettura piramidale: sia in entrata (recettore) che in uscita (effettore) lo stesso campo distrettuale è rappresentato più volte e su piani sovrapposti ove ciascun piano è espressione simbolica di un dominio spazio-temporale. La mole dei flussi informazionali sottostanti è articolata nello spazio/tempo per sottrazioni discrete e incessanti quasi come avverrebbe nella creazione di una sinfonia ottenuta inserendo ordinate pause di silenzio in un caos di rumori assordanti. In ogni caso riconoscere al SN il ruolo di “trasportatore verso l’alto” del flusso informazionale contrattile non aiuta molto a chiarire alcuni aspetti del destino informazionale centrale del flusso biochimico. Abbiamo citato la presenza di interfacce integrativo/elaborative tra il livello nervoso pre-spinale (il Sistema Nevoso Vegetativo –SNV– centralizzato nei gangli pre- e laterovertebrali) e la superficie recettoriale linfocitaria. La mole di tale flusso informazionale è articolata secondo un dominio spazio-temporale scandito da fenomeni di coordinazione/inibizione gestiti da questo stesso livello integrativo nervoso. La colica intestinale ne rappresenta un esempio efficace in Patologia: all’arresto della peristalsi si associa la contrattura trasversa del lume in concomitanza con l’attivazione linfogranulocitaria locale conseguente a uno stimolo irritativo mucoso. La perdita della fisiologica coordinazione dei movimenti che caratterizzano l’incessante e armoniosa motilità gastrointestinale è attivamente indotta dall’azione dei gangli periviscerali del parasimpatico cranio-sacrale e dai gangli paravertebrali del simpatico. L’impossibilità di sciogliere intenzionalmente il nodo doloroso della colica mostra chiaramente come, specie in Patologia, si disveli il reale significato del termine “Autonomo”, caro agli autori anglosassoni, con cui il SNV spesso viene indicato. Tuttavia con ogni probabilità anche questo traffico informazionale viene “simbolificato” e centralizzato attraverso le vie afferenti ganglio-spinali. Il problema è che l’inibizione della competenza immunitaria passa attraverso il fenomeno della tolleranza linfocitaria. Quest’ultima trova un punto di regolazione tramite leve funzionali attivate dal flusso informazionale nervoso ma di cui non abbiamo un quadro preciso. Da oltre trent’anni tra le leve regolatorie del traffico di espressione linfocitaria è stato individuato il sistema degli interferon, famiglia di molecole considerata il più potente modulatore della tolleranza linfocitaria [31] [48] [58]. Non si ha notizia di capacità secretive di interferon da parte di strutture gangliari autonomiche. Tuttavia queste ultime esprimono adenosina e purina, sostanze in grado di modulare la sensibilità recettoriale dei linfociti e dei monociti attraverso il sistema degli interferon e questo potrebbe rappresentare l’indizio di una regolazione indiretta. Ben più note sono le proprietà neuromodulatorie centrali esercitate dagli interferon su sistemi apparentemente estranei alla regolazione immunitaria come le aree catecolaminergiche e serotoninergiche mesiali dell’encefalo: dagli anni Novanta del XX° Secolo fino a oggi si è sviluppata una vasta bibliografia sugli effetti psichiatrici indesiderati in pazienti affetti da sclerosi multipla trattati a lungo con beta-interferon [40]. Quanto fin qui esposto costituisce un modello interpretativo fisiologico di realtà anatomico-funzionali già note. Proveremo ora a cimentarlo con alcuni fenomeni correlati ad alcune patologie.
30d.
Il conflitto tra dominii spazio-temporali. Modelli patogenetici oltre l’Omeostasia di Cannon
“Il sistema nervoso interagisce con l’ambiente che lo circonda modificando incessantemente la propria struttura… cosicchè fatalmente esso si trova a esprimere, in ogni momento, la registrazione di tutti i propri pregressi cambiamenti strutturali…. è come se, mentre elabora le informazioni che gli provengono dall’ambiente, esso generasse un mondo interno proprio” (Fritjof Capra).
Ogni bioritmo è perturbabile da qualsiasi leva funzionale operante da qualsiasi settore somatico pure se remotamente connesso. E’ proprio qui che si trova la base fisiopatologica della “noxa”, cioè di ogni evento scaturito dall’interazione tra organismo e ambiente che induca perturbazioni su un bioritmo attraversandone un flusso informazionale. Il bioritmo perturbato verrà immediatamente e inevitabilmente rappresentato su tutto il suo dominio spazio-temporale attivando tutte le leve funzionali a cui è connesso con una progressione “a domino”. Il processo dell’infiammazione è una perfetta esemplificazione di questa modalità di espansione: lo scenario suscita l’immagine di una polveriera pronta a esplodere non appena si accenda una fiamma ma, in realtà, tutta l’interazione con l’ambiente può essere interpretata come una sequenza di “accensioni” potenzialmente drammatiche. A impedire le conseguenze più gravi non è tanto la mancata attivazione di processi dirompenti quanto piuttosto la capacità di spegnimento dei focolai alla loro prima insorgenza espressa dai dispositivi di coordinazione/inibizione. Anzitutto sulla funzionalità di questi ultimi è logico perciò che si orienti l’attenzione di chi osserva il dispiegarsi dei fenomeni patologici immunomediati.
Per definire correttamente un “dominio spazio-temporale” occorre anzitutto individuarne la ritmologia: fenomeni che si innescano, si protraggono e si esauriscono nell’arco di anni-mesi, giorni-ore o minuti-secondi esprimono flussi informazionali e leve funzionali cadenzate su altrettante scansioni temporali. Generalmente tanto più lungo è il periodo in cui il ritmo si esprime quanto più filogeneticamente antichi sono i dispositivi implicati e tanto maggiore è la loro estensione spaziale e il grado di connettività tra diversi flussi informazionali. Per individuare i bioritmi di anni-mesi vanno ricercati funzionalmente gli effettori dei processi differenziativi tessutali, anzitutto i Fattori di Crescita, le cui relazioni con il Sistema Immunitario (SI) appaiono sempre più rilevanti: è provato infatti che essi dettino le proprietà funzionali e il fenotipo dei macrofagi e delle cellule T. Oltre a questo è rimarchevole che siano state rilevate in corso di processi infiammatori attività protettive espresse dall’interazione tra Fattori di Crescita e citochine [55][56]. Il ruolo della coordinazione/inibizione è generalmente più apprezzabile nel contesto dei fenomeni che si stagliano in un ritmo ultracircadiano. Consideriamo un episodio influenzale: qualunque sia stato il veicolo d’ingresso la via informazionale biochimica (ossia nutrizionale/immunitaria) porta con sé dei determinanti antigenici (virus o sue parti) attraverso un’interfaccia più o meno liquida fino al sito di integrazione/elaborazione ove incontra la cellula interprete. Questa, dotata di recettorialità biochimicamente competente (diversi tipi di cellule in relazione a diversi gradi di liquidità dell’interfaccia) produce leve funzionali (le citochine) che sollecitano a raggiera vie informazionali convergenti sui successivi siti di integrazione/elaborazione. Alcune di queste vie saranno situate su un livello di più alta risoluzione spazio-temporale interpretato da flussi informazionali ancora per lo più biochimici. L’attivazione immunospecifica linfocitomediata emerge infatti a distanza di tempo dall’incontro tra l’antigene e l’anticorpo e in quel compartimento segregato rappresentato dal linfonodo. A seguito della perturbazione altre vie percorse saranno quelle pertinenti al flusso contrattile elettrochimico (ovvero “nervoso”) ed è proprio quest’ultimo che nel quadro clinico assumerà alla fine la maggiore visibilità. In corso di sindrome influenzale assistiamo infatti a iperreflessia cutaneo-muscolare diffusa associata a riduzione della coordinazione dei movimenti intenzionali, disordini della motilità gastrointestinale, emergenza di iperpatia, dispercezioni sensoriali, dolori, ipersensibilità mucoso-cutanea e febbre. Tutti questi fenomeni corrispondono di fatto a una moderata e diffusa riduzione della segregazione funzionale dermato-metamerica con disgregazione dei campi recettoriali coscienti. Per quanto riguarda la febbre sono degne di nota le interpretazioni teleologiche che ha ispirato: invocando una vulnerabilità dei virus alle temperature elevate si è giunti a sostenere che, omeostaticamente, il rialzo termico corporeo corrispondesse a un dispositivo antivirale [6]. Se l’ipotesi lascia perplessi (le temperature febbrili favoriscono la crescita di molti microorganismi piuttosto che inibirla) il nostro modello può offrire una tesi più solida: la sindrome influenzale non esprimerebbe semplicemente la risposta di un sistema colpito da un insulto bensì l’emergenza dal basso di un flusso informazionale appartenente a un dominio spazio-temporale sottostante. Inoltre il pirogeno endogeno prodotto dai leucociti non induce una condizione di mero e persistente incremento della temperatura corporea: diversamente dall’ipertermia la febbre qui consiste nello slittamento delle normali risposte neurovegetative rispetto alle sollecitazioni termiche ambientali come se fosse stata inserita una “tara” a perturbare le funzioni di una bilancia: la meccanica della bilancia continua a funzionare ma i valori espressi risultano esorbitanti rispetto all’usuale regime di variabilità. Sotto questa luce la tesi della febbre quale antidoto termico appare assai debole. Se invece pensiamo a un regime ancestrale, a un contesto individuo-ambiente semplificato e originariamente immerso in temperature più alte, possiamo considerare un’ipotesi che giustifica anche le altre manifestazioni influenzali: attivato dall’interazione antigene-anticorpo un flusso informazionale nervoso più antico emerge occasionalmente come principale regolatore della funzionalità globale venendo a porsi in una dimensione spazio-temporale attigua a quella del flusso biochimico attivante. Al sistema attivato la presenza del virus “non interessa”: esso semplicemente manifesta un modello nervoso-comportamentale ancestrale. Anche tutte le altre caratteristiche dell’emergente flusso informazionale nervoso (dalla disestesia alle manifestazioni viscero-somatiche) testimoniano la contiguità con il tipo di flusso biochimico attivato dall’interazione con l’ambiente. Rispetto alla classica visione della malattia quale espressione di un danno che mette in crisi l’equilibrio omeostatico tra leve funzionali, qui emerge un modello interpretativo diverso.
Un elemento comune al funzionamento del SN e del SI che si staglia con tutta evidenza è la “memoria”. Una buona definizione di memoria, che sia comprensiva tanto dei fenomeni osservabili in strutture biologiche semplici quanto nella manifestazione di facoltà mentali complesse, è quella di modulazione dell’entità di una risposta in relazione alla reiterazione di uno stimolo sempre uguale [3]. In ambito neurofisiologico il più elementare modello di memoria è rappresentato dalla variazione di soglia di eccitazione di una membrana cellulare in relazione a una sequenza di stimoli elettrici di uguale intensità. Tale proprietà è detta “accomodazione di soglia di membrana”. Siamo abituati a considerare membrane “eccitabili” solo quelle che, stimolate, danno luogo a perturbazioni di voltaggio sovramassimali detti “Potenziali di Azione” (PdA) o “spike”. Si tratta delle membrane che realizzano i flussi informazionali elettrochimici nei muscoli lisci, nei muscoli striati e nei neuroni. In realtà i meccanismi molecolari che sovraintendono l’eccitabilità sono presenti (e in buona parte analoghi) in tutti i tipi cellulari. Essi costituiscono i fondamentali sistemi di trasporto intracitoplasmatico di sostanze e di movimento di singole cellule in diversi compartimenti tessutali (diapedesi leucocitaria, movimento degli spermatozoi, eccetera). E’ perciò evidente che anche nei flussi informazionali nutrizionali-immunitari si riscontrino fenomeni di modulazione dell’eccitabilità. In buona sostanza sono stati assegnati nomi differenti a oggetti che invece sono tra loro molto simili: la modulazione dell’eccitabilità di un neurone non è frutto di processi molto diversi da quelli che regolano la sensibilità recettoriale di un linfocito o di una cellula tiroidea. Piuttosto che differenze qualitative tra queste manifestazioni sembrano sussistere differenze quantitative poiché riflettono le differenti risoluzioni spazio-temporali in cui viaggia ciascun flusso informazionale.
Torniamo al modello fisiopatologico. Dato che una perturbazione indotta da un’interazione ambiente-individuo comporta il dispiegamento di flussi informazionali convergenti su un determinato dominio spazio-temporale c’è anzitutto da chiedersi quale sia la soglia di stimolo necessaria e sufficiente a promuovere tale dispiegamento e quali siano i fenomeni adattativi che l’evento sollecita (sempre che esso non provochi danni biologici irreversibili). L’introduzione del concetto di “soglia variabile” (11b) anzitutto scardina l’idea riduzionistica per cui un evento patologico debba manifestarsi in forma tanto più grave quanto più intenso sia stato lo stimolo: specie nelle malattie croniche i rapporti dimensionali tra agente etiologico e manifestazioni patologiche possono difettare di ogni proporzionalità ed è questo il motivo per cui spesso risulta impossibile identificare l’agente etiologico. Entra qui in gioco la cellula che funge da interprete nel luogo di interazione tra flussi informazionali neuro-contrattile e nutrizionale-immunitario, cioè nel teatro del processo di integrazione/elaborazione. Dal momento che in risposta alla noxa l’interprete produce la “leva funzionale” risulta evidente la sua competenza nei confronti dell’interazione. Il suo compito si estende però anche alla modulazione di soglia: memore dei precedenti eventi informazionali su cui era stabilito e ogni volta in misura diversificata, l’interprete si rende testimone dell’entità della leva funzionale da esprimere. Tale variabilità è alla base di fenomeni clinici noti in Immunologia nella forma delle manifestazioni che spaziano dalla tachifilassi delle allergie alla tolleranza immunitaria (“immunizzazione”, vaccinazione). L’acquisizione della variabilità di soglia da parte dell’interprete non si esprime in una singola sede di integrazione/elaborazione ma si rinnova in tutti gli altri passaggi integrativi percorsi dai flussi informazionali, cioè tanto sulla linea neuro-contrattile quanto su quella nutrizionale/immunitaria. Quando una perturbazione si espande nel suo dominio spazio-temporale ogni punto di interazione integrativo/elaborativa diviene sede di competenze mnesico-adattative venendosi così a creare nel reticolo spazio-temporale del flusso informazionale una “accentuazione del solco” su particolari percorsi. Se al termine del ciclo perturbativo le strutture biologiche restano sufficientemente integre le accentuazioni del solco vengono ricoperte dal ritorno del bioritmo precedente (cioè dalle competenze di coordinazione-inibizione esercitate dai domini di flusso informazionale sovrastanti). In questo caso apparentemente non resta nulla della perturbazione appena passata visto che il ripristino dell’ordine gerarchico sarà stato in grado di coordinare/inibire nuovamente i flussi informazionali secondo le scansioni spazio-temporali antecedenti. Ma se un’ulteriore perturbazione riproporrà l’influsso di un dominio spazio-temporale più basso potranno riemergere i vecchi “solchi mnesici” di percorso già tracciati entro il reticolo informazionale. Queste nozioni, oggi corroborate da robuste evidenze neuroimmunologiche, vennero dimostrate sperimentalmente già nella terza decade del XX° Secolo dal geniale ricercatore sovietico Alexandr Dmitrievich Speranskij (1988-1961). Egli osservò a lungo quanto accadeva quando sugli animali del suo stabulario provocava lesioni sperimentali distanziate nel tempo che lui identificava come irritazioni nervose. Dopo la prima lesione alcuni modelli animali sembravano non patire alcuna conseguenza mentre in altri poté osservare la comparsa di quadri morbosi diversi (alopecie, ulcere della mucosa enterica, emorragie polmonari, edemi, cheratiti, papillomi, eccetera) che, entro tempi variabili, recedevano con apparente guarigione dell’animale. Generalmente negli animali che rispondevano alla noxa più periferico era stato il distretto somatico oggetto della prima lesione sperimentale più lunga si mostrava la latenza temporale tra questa e la comparsa dei sintomi. Nel primo gruppo, cioè nei modelli animali apparentemente non reattivi, la ripetizione dell’irritazione/lesione sperimentale poteva determinare la comparsa di uno dei quadri clinici descritti in forma drammatica (talora persino letale) mentre nel secondo, quello dei soggetti apparentemente guariti, il quadro clinico che si era manifestato in forma leggera ora si replicava anche qui in forma severa e minacciosa per la vita. Speranskij ne concluse che doveva esservi una sorta di “memoria per il tipo di irritazione” e che per la manifestazione dei quadri morbosi non fosse importante la natura dello stimolo irritativo quanto piuttosto l’irritazione stessa e la conseguente risposta nervosa: “Il processo neurodistrofico prodottosi nel sistema nervoso a seguito della lesione sperimentale può fermarsi senza lasciare traccia di sé. Talvolta però esso permane in forma latente pronto a riemergere a seguito di uno stimolo qualsiasi. In questi casi si può assistere alla replica della reazione corrispondente al processo che già era stato osservato in precedenza” [47].
In sintesi, l’interazione con l’ambiente è fonte continua di lesioni/irritazioni e di alterazioni cito-istologiche indotte da stimoli fisico-chimici. Rispetto alle relazioni meccaniche osservabili nella materia inorganica l’evoluzione delle specie biologiche mostra qualcosa di più complesso: nel vivente la presenza della noxa quale elemento costante costituisce veicolo di apprendimento tramite “solchi mnesici” che modellano le caratteristiche dell’adattamento individuale (potremmo quasi definirlo “culturale”). Le tracce mnesiche sviluppate nell’individuo in esito alle sue relazioni con l’ambiente nella fase di riproduzione della specie vengono inserite nelle sue caratteristiche genetiche. L’epigenetica, scienza moderna assai feconda, descrive esattamente questi processi.
30e.
Le malattie reumatiche: la complessità compressa nell’angustia della “medicina d’organo”
Agli albori della Medicina il termine “reuma” ha indicato un flusso di malattia che accende l’infiammazione in sedi somatiche diverse per struttura e funzione: cuore, polmoni, cervello, reni, intestino e (soprattutto) vasi, articolazioni e cute. Questa vastissima categoria nosologica dalla non facile sistematizzazione [17] sembra costituire per la sua incidenza la patologia cronica peculiare di quest’epoca rappresentando il maggiore capitolo di spesa sanitaria e la maggiore causa di inabilità motoria. Da un lato vi troviamo patologie altamente invalidanti (l’artrite reumatoide, il lupus eritematoso sistemico, eccetera) e dall’altro condizioni che spesso vengono considerate quali normali malanni dell’età avanzata (artrosi, spondilodiscoartrosi, eccetera). Nello studio di questa categoria di malattie la scienza di base è l’Immunologia che qui, diversamente da quanto accade per le altre specialità, sconta l’impossibilità di definire un ambito settoriale basato sui criteri della Patologia d’Organo. Eppure, secondo l’impostazione filo-ontogenetica che qui proponiamo, in queste malattie si rende ben identificabile l’organo bersaglio dato che il “flusso malefico” scorre in quella dimensione spazio-temporale facilmente identificabile nello stroma. Rammentiamo che esso rappresenta quel mezzo semifluido reticololamellare che individuammo nello sviluppo delle vie biochimiche nutrizionali/immunitarie e nel cui ambito il ruolo di cellula interprete è affidato a elementi ematolinfopoietici. Ma la localizzazione dell’ambito in cui si verificano le perturbazioni patogenetiche, ossia lo stroma, non basta da sola a definire i fenomeni fisiopatologici alla base del “reuma”. Reinterpretare i fenomeni secondo l’impostazione metodologica da noi adottata richiede invece la soluzione di almeno quattro quesiti: su quale dimensione spaziale si stagliano queste perturbazioni? In quale dimensione temporale si snodano? A quale soglia di eccitabilità si collocano i flussi informazionali rispetto alle loro “nuove” sedi di integrazione/elaborazione? E infine, last but not least, come si articolano i sistemi di controllo mediante coordinazione/inibizione su questi nuovi “dominii patologici”? Al termine di questa disamina si potrebbe assistere a un cambiamento di ruolo dello stesso stroma: da target a “primo attore” delle patologie reumatiche.
Avendo prospettato l’esistenza di un flusso informazionale neuro-contrattile quale attore delle manifestazioni cliniche delle malattie (che si esprime attraverso la disgregazione dei sovrastanti dominii coordinativi/inibitori) riconsideriamo le caratteristiche delle immunopatie attraverso l’analisi del flusso integrato sul dominio spazio-temporale ormonale: l’asse ipotalamo-ipofisario. Non enumeriamo qui le relazioni, note da tempo, tra detto asse e la modulazione della risposta immune. La ricerca biomolecolare scopre sempre nuovi intrecci interattivi tra effettori e recettori arricchendo esponenzialmente il pleiomorfismo funzionale di secreti da parte di nervi, cellule (stanziali o circolanti) e ghiandole endocrine scardinando così, pezzo a pezzo, la vecchia concezione che vedeva separati i sistemi Nervoso, Endocrino e Immunitario. Una delle relazioni più recenti concerne l’attivazione del Nerve Growth Factor (NGF), fattore di crescita e differenziazione embrionale secreto da cellule stanziali (i mastociti) in risposta al danno tessutale. Questi ultimi, a loro volta, promuovono fenomeni di “sensitizzazione nervosa” (31) responsabili del dolore neuropatico locale [1][28][29][38]. In questa sede occorre anzitutto conseguire una visione d’insieme evitando che i meccanismi molecolari si perdano in un caotico calderone senza logica. Il dominio spazio-temporale rappresentato da questo livello di flusso informazionale gestisce a un tempo istanze nutrizionali, riproduttive e comunicative. I tempi di scansione abbracciano periodi variabili con fenomeni che si estendono su scala multicircadiana breve (pensiamo ad esempio ai ritmi secretori dell’ossitocina) passando da quelli che descrivono cicli mensili con andamento pulsatile fino a quelli che si svolgono nell’arco di anni (come le dinamiche biologiche retrostanti alla pubertà). Le “leve funzionali” mostrano chiaramente la molteplicità complanare dei flussi informazionali su cui si attivano le risposte secretorie e motorie: da quelle tipicamente sinaptiche (rapide e localizzate) a quelle dispiegate attraverso il flusso circolatorio e propriamente definite endocrine (lente e diffuse). Sono tutte attivazioni “sinergiche” capaci di innescare modalità di funzionamento dell’intero organismo in cui ciascun elemento risponde secondo un intrinseco concerto di effetto con gli altri (sympathòs). Da questo punto di vista la distinzione tra Sistemi Nervoso ed Endocrino ha valore solo se è riferita alle diverse caratteristiche strutturali dei due sistemi e non da quello del risultato funzionale. L’ambito spaziale della patogenesi reumatica parte da un flusso informazionale di tipo nutrizionale/immunitario di livello elevato, cioè già ampiamente processato (un complesso antigene-anticorpo, un Antigen Presenting Cell, una cellula infettata da virus o che esibisce caratteristiche pretumorali). Si tratta di un soggetto già sotto l’egida di un SI circoscritto ed elaborato in un’architettura reticololamellare che potremmo definire “complesso informazionale integrato”. Esso accede a vie di comunicazione selettive ed embricate capaci a un tempo di scorrere lentamente attraverso le vie stromatico-reticolari che lo ospitano ma, a tratti, anche di fluire assai più rapidamente attraverso le ben più liquide vie linfo-ematiche quando i prodotti delle loro interazioni si liberano negli spazi intercellulari. Le “leve funzionali” scaturite dalle interazioni di questo nuovo flusso informazionale saranno modulate, quanto a potenza, diffusione e durata d’azione in relazione al dominio spazio-temporale in cui avviene l’interazione tra i flussi. Pensiamo a un estremo rappresentato da una crisi asmatica (ove le interazioni avvengono tra flussi informazionali che viaggiano su un dominio spazio-temporale rapido e relativamente circoscritto) a fronte di un estremo opposto come quello rappresentato da una poliartrite cronica. Nel primo caso le leve funzionali attivate saranno prevalentemente tarate su effetti brevi, potenti e circoscritti mentre nel secondo saranno coinvolti meccanismi di attivazione prolungati e ad ampia estensione topografica. L’attivazione delle “leve funzionali” promuove così fenomeni di attivazione infiammatoria locale che si sommano con segnali che accendono periferie remote in base a meccanismi interpretativi localizzati capricciosamente. Nel dominio d’osservazione temporale emerge un altro fenomeno: gli eventi perturbativi sembrano serbare in sé stessi diverse potenze di erogazione in relazione a eventi legati ad altri flussi informazionali. La letteratura scientifica e una sconfinata empirìa offrono argomenti sulle relazioni tra riaccensioni acute in corso di malattie infiammatorie croniche e svariati eventi interattivi (dallo stress psicofisico al periodo mestruale, dalle variazioni metereologiche ai regimi alimentari). Insieme alla variazione della “potenza d’erogazione” delle leve funzionali (citochine infiammatorie, fattori d’attivazione mastocitari e piastrinici) la variazione nel dominio temporale del flusso nutrizionale/immunitario si esibisce nella sua caratteristica intermittenza: pur non venendo meno l’istanza d’accensione, che il più delle volte rimane indovata e misconosciuta, il flusso perturbante si accende e si spegne autonomamente secondo una ritmologia ultracircadiana, talora ultramensile quando non addirittura annuale. Dal punto di osservazione del clinico la precipua ritmologia delle patologie immunologiche croniche è un dato talmente ricorrente da diventare un forte indizio diagnostico. Interpretando questo nell’ambito del percorso filo-ontogenetico che abbiamo abbozzato il bioritmo che caratterizza questi stati patologici mostra alcuni elementi pulsatili che ritroviamo in un’istanza comportamentale successiva e complementare alla nutrizione: la sessualità. Non siamo più dinanzi a un organismo pluricellulare in competizione solo per i nutrienti ma adesso anche per la ricerca del partner entro la propria specie e possiamo pensare, in termini di “leva funzionale”, a certe attività vasomotorie connesse a manifestazioni comunicative in alcune specie nella fase di corteggiamento: l’attivazione del flusso dermico in precise parti del corpo per produrre effetti di variazione cromatica o di deformazione volumetrica, variazioni termiche e (per via esocrina) produzione di emanazioni olfattive. Sul piano spaziale queste localizzazioni corporee variano da specie a specie secondo disposizioni che non sempre alludono all’atto copulativo: esse possono enfatizzare la dimensione somatica, rappresentare capacità aggressive (agilità, energia muscolare), talora semplicemente esibire l’armamentario cromatico a riprova della buona fruizione individuale del patrimonio genetico (pensiamo al pavone). Si tratta di una riserva simbolica che recluta dispositivi funzionali secondo criteri specie-specifici dislocati su strutture eterogenee. Il ritmo delle perturbazioni assume caratteristiche pulsatili (“bursts”: scoppi) e nell’arco di qualche settimana si sviluppano eventi a catena che mimano l’accensione progressiva di fuochi d’artificio a cui segue una pausa e poi una nuova accensione. Presso i mammiferi inferiori i bursts assumono ritmologie nell’ambito dei mesi lunari e, su una dimensione più lunga, degli equinozi: qui riconosciamo gli estri, i periodi dell’amore. Questi ritmi, in relazione alla durata della vita, realizzano perturbazioni che si susseguono come grappoli di eventi ravvicinati in una distribuzione temporale globale sinusoidale: a una fase silente segue una successione crescente di eventi moltiplicativi che raggiungono un apice, poi una deflessione parziale, poi un altro apice, poi una successiva depressione più marcata e infine una sequela di incrementi via via più deboli fino a una lenta discesa verso i valori basali. In vecchiaia il ritmo sinusoidale, fino ad allora sessualmente guidato, diventa inconsistente e si torna alla condizione nutrizionale di fondo.
Tornando all’origine del complesso informazionale integrato conosciamo una serie di interazioni immunitarie formulabili in quattro modelli patogenetici: la reazione crociata antigene-anticorpo tra antigene self e non-self [10], la via di attivazione del complemento [37], la formazione di immunocomplessi [41] e la sensibilizzazione linfocitaria [2]. Nelle malattie disimmuni queste modalità interattive spiegano dei passaggi più o meno riconoscibili ma non sempre in modo lineare nelle diverse patologie e con manifestazioni che non sempre le rendono tra loro nettamente distinguibili. Quest’incertezza proviene dal fatto che non è possibile attribuire univocamente un dato meccanismo a una precisa patologia e perché tutti i suddetti modelli patogenetici difettano di un chiaro significato di primum movens. Sappiamo infatti che la sensibilizzazione linfocitaria può preludere alla secrezione di un clone di immunoglobuline capaci a loro volta di attivare fenomeni di reattività crociata o che l’attivazione di una cascata complementare da parte di un epitopo antigenico può indurre una sensibilizzazione linfocitaria rivolta ad altri epitopi. Al proposito il geniale immunologo Niels Kaj Jerne nel 1974 riuscì a descrivere questo paradigma universale [24]. Sappiamo anche che la produzione di immunoglobuline, in qualsiasi modo suscitata, può indurre la formazione di immunocomplessi circolanti o infiltranti lo stroma. Nonostante la potenziale multidirezionalità di questi fenomeni possiamo riconoscere molte malattie proprio dal tipo di fenomeno prevalentemente innescato. Ecco che si ripropone il tema dell’accentuazione del solco mnesico: il “combinato disposto” tra un solco mnesico trasmesso geneticamente, ulteriori solchi mnesici indotti dalle prime interazioni nutrizionali/immunitarie (in fase intra- ed extrauterina) e un successivo più o meno traumatico percorso conoscitivo individuale esitano in innumerevoli combinazioni relazionali tra individuo e ambiente e tra settori dello stesso organismo che si situano su sedi di elaborazione/integrazione relativamente ricorrenti. E’ frequente che i pazienti raccontino che la malattia reumatica è iniziata dopo una gravidanza o che è peggiorata dopo un’influenza, un trauma o un intervento chirurgico. Questa ricorrenza di scambio informazionale su medesimi punti cruciali del flusso informazionale nutrizionale-immunitario è alla base dell’epidemiologia di tante “misteriose” malattie sempre a cavallo tra indizi di predisposizione genetica e influssi geografico-ambientali come è il caso della distribuzione geografica della Sclerosi Multipla [44][26].
Al di là delle localizzazioni primarie che caratterizzano diverse immunopatie (quindi al di là dell’identità specifica del complesso informazionale integrato) è utile focalizzare due aspetti derivanti dall’elaborazione/integrazione: il primo è l’espressione delle “leve funzionali” che derivano da ciascuna interazione, il secondo è la regolazione di soglia che tali interazioni producono sui flussi informazionali sovrastanti. Entrambi vanno visti nell’ambito dei dominii spazio-temporali che vanno a occupare. Per quanto riguarda i fenomeni di erogazione di una leva funzionale da parte di cellule interpreti è nota la secrezione di “chitochine ubiquitarie” da parte di cellule più o meno inserite nel flusso informazionale nutrizionale/immunitario: la triade TNF, IL2 e IL6 [9]. Attraverso vie privilegiate il complesso informazionale integrato viaggia in compartimenti specifici e comunicanti e diventa in grado di attivare cellule interpreti attraverso strutture recettoriali sensibili al contenuto informazionale stesso. Ciò può avvenire in virtù di una perturbazione limitata entro la sua stessa capacità di auto-contenimento ovvero in virtù di un successivo deragliamento di funzione destinato a dare luogo a fenomeni distruttivo-ricostruttivi successivi. Nel primo caso queste “leve funzionali” attivano, su domini spazio-temporali complanari, cascate di risposta ormonale e nervosa. Le risposte ormonali spaziano dai Fattori di Crescita Embrionali ai secreti dell’asse ipotalamo-ipofisario mentre tra i fattori nervosi sono molto rappresentati quelli attivati dalle cellule astrocitarie integrate nei gangli dorsali spinali. Nel secondo caso, quando siano attivati fenomeni di distruzione-ricostruzione, entrano in gioco da un lato gli stessi Fattori di Crescita Embrionali induttori di rimodellamento strutturale dei tessuti [20][32][39][46] e dall’altro eventi di adattamento recettoriale nervoso che modulano la soglia di eccitazione locoregionale [21][45].
Quanto all’andamento temporale nelle patologie disimmuni osserviamo tipicamente eventi pulsatili ultra-circadiani organizzati in fasi di accensione-spegnimento dell’infiammazione: le pousseés. Nei dominii regolatori di livello ipotalamo-ipofisario le connessioni tra attività nervose e influenze endocrine appaiono talmente numerose e pervasive che si rischia di identificare il Sistema Neuroendocrino quale unico gestore dell’espressione fenotipica delle immunopatie. Infatti, a proposito dei dominii regolatori attinenti alla sfera riproduttiva, in queste malattie sono evidenti la forte preponderanza di genere, le relazioni temporali delle recrudescenze con i cicli mensili e la storia naturale che spesso coincide col ciclo di maturazione-esaurimento della capacità riproduttiva. Ma tale semplificazione liquiderebbe la realtà di malattie altrettanto “disimmuni” con opposte predisposizioni di genere: pensiamo alla spondilite anchilosante che colpisce prevalentemente soggetti di sesso maschile rispetto all’artrite reumatoide che predilige il sesso femminile [12] e il fatto che nella stessa malattia possono verificarsi andamenti assai eterogenei. L’andamento delle malattie infiammatorie croniche offre quasi sempre a considerare quadri progressivo-continui, quadri a decorso variabile prima con relapsing-remitting e poi con andamento progressivo-continuo e, infine, quadri monofasici episodici. La stessa ipervariabilità si riscontra nella distribuzione topografica di queste patologie. Pertanto più che a un dominus neuro-endocrino sembra più logico pensare a influenze “a geometria variabile” in cui, a parità di accensione infiammatoria da parte dello stroma, il flusso informazionale neuro-endocrino si esprime in misura distinta e circostanziata imbattendosi via via con diverse soglie di eccitabilità inserite nei diversi solchi mnesici che percorre (esiti questi della storia filo-ontogenetica individuale).
30f.
Brevi richiami di carattere anatomo-fisiologico
All’inizio della trattazione ci siamo soffermati sul principio per cui la perturbazione di un bioritmo può rappresentare il risultato di un’elaborazione-interazione prodotta lungo il percorso di qualsiasi flusso informazionale purché sussistano vie di comunicazione percorribili dalle leve funzionali innescate. Abbiamo esposto come le vie di comunicazione siano garantite da due modalità di propagazione: quella nutrizionale-immunitaria (che scorre entro spazi a maggiore o minore densità stromale) e quella neuro-contrattile (che scorre in percorsi di accoppiamento elettrochimico tra sistemi contigui). In realtà nel Sistema Nervoso Centrale (SNC) le due modalità di propagazione del flusso informazionale si unificano integrandosi secondo un’ulteriore evoluzione architettonica: alla contiguità “sinciziale” tra cellule si sovrappone una dinamica di allungamento-differenziazione della cellula nervosa che assume così una polarità tra un compartimento afferente (dendrita) e uno efferente (assone). Entrambi i poli contraggono poi una comunicazione di flusso di nutrizionale/immunitario con altre cellule nervose nel compartimento delimitato ed esclusivo dello spazio sinaptico. Tali strategie evolutive rispecchiano istanze individuate nel sistema pluricellulare dell’Hydra e nella disposizione anatomo-funzionale dell’intestino (elongazione, avvoltolamento, cefalizzazione).
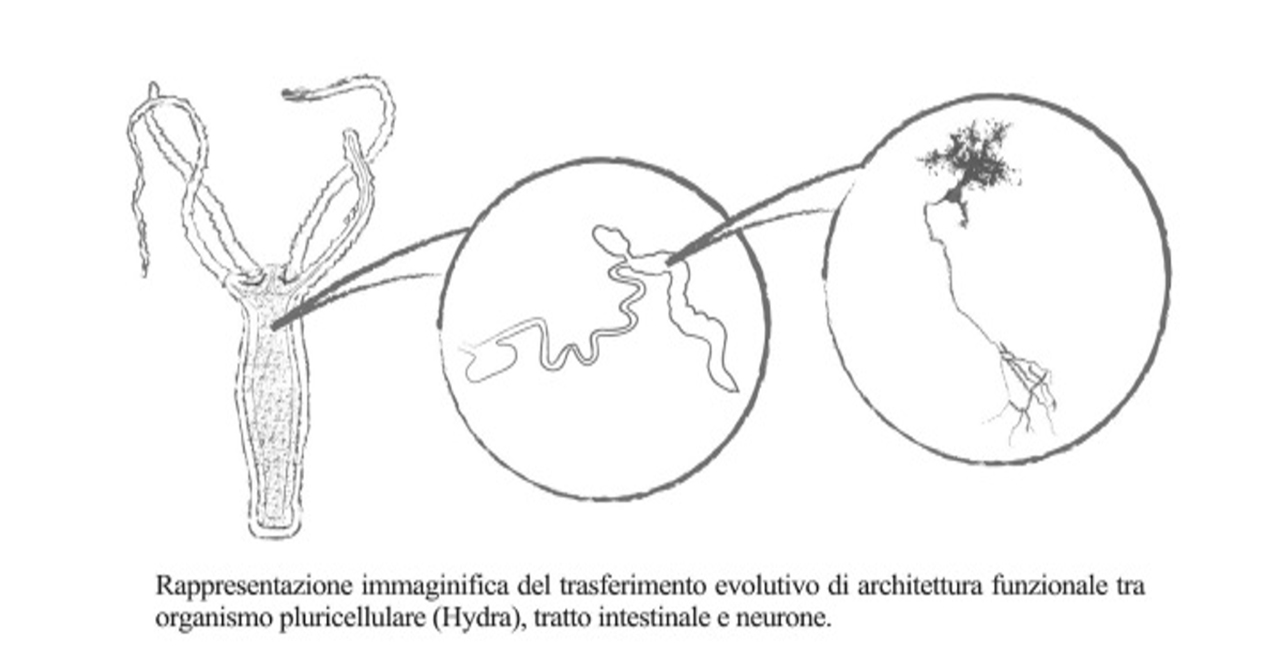
Nella produzione informazionale nervosa sia i fenomeni contrattili del versante elettrochimico che i costituenti nutrizionali/immunitari del versante biochimico sono occultati e simbolificati: della perturbazione contrattile viene trasmessa solo la concomitante variazione di potenziale elettrico di membrana mentre della perturbazione nutrizionale/immunitaria viene sintetizzata e impiegata solo una molecola (il neurotrasmettitore sinaptico) di per sé priva di alcuna significativa capacità nutrizionale. L’uso di simboli semplificati permette una notevole economia di gestione del materiale biochimico purché i flussi informazionali vengano gestiti all’interno di una rigida segregazione architettonica che garantisca l’univocità di significato dell’informazione trasmessa.
Guardiamo ora più addentro al flusso informazionale nervoso, centralizzato nella cellula nervosa e nei suoi contatti sinaptici. Nel SNC tutte le istanze della periferia devono essere rappresentate e il “lavoro informazionale” dell’intero sistema deve disporre di tutti i gradi di risoluzione spazio-temporale su cui si staglia la relazione individuo/ambiente. Peraltro il SN non si occupa solo delle relazioni nutrizionali/immunitarie: infatti l’interazione con l’ambiente si esprime soprattutto attraverso capacità locomotorie e comunicative. Per quanto riguarda queste ultime l’interfaccia con l’ambiente si avvale di interpreti (aree di membrana pre- e postsinaptiche) che si dispongono in superficie (cute in afferenza, muscoli e ghiandole in efferenza) secondo l’architettura già descritta per riconoscere la complanarietà dei dominii spazio-temporali: in aree ampie, scarsamente demarcate e con bassa risoluzione spazio-temporale troviamo intermediari di eventi lenti (temperatura, variazioni delle concentrazioni chimiche, eccetera) mentre entro aree puntiformi con più alta risoluzione spazio-temporale troviamo interpreti di eventi rapidi (pressione, tensione, passaggi di contrasto). Molteplici flussi informazionali formatisi dall’interazione con l’ambiente tendono a raccogliersi in distinti “cavi” di conduzione, ciascuno rappresentativo del proprio dominio spazio-temporale. Struttura, composizione e percorso di ciascun cavo riflettono il destino di ciascun flusso entro l’architettura globale del SN. Qualcosa di simile avviene quando raccogliamo tutti insieme in un portacavi i fili del nostro personal computer: pur facendo questa operazione a caso (cioè senza badare al calibro e alla funzione dei singoli fili) le giuste connessioni con le “periferiche” non risultano pregiudicate. Per correttezza terminologica i “portacavi” di informazione tra ambiente esterno e le stazioni nervose centrali più prossime alla periferia si definiscono nervi mentre quelli che viaggiano nel SNC fasci. Nell’ambito del nervo distinguiamo ciascun assone in relazione diretta con il tipo di dominio spazio-temporale dell’informazione che trasmette: le vie informazionali a bassa risoluzione spazio-temporale sono rappresentati da assoni di piccolo calibro e privi di guaina mielinica mentre quelle a maggiore risoluzione spazio-temporale sono rappresentati da assoni di calibro via via più largo con tratti sempre più lunghi di rivestimento mielinico (cellule di Schwann). Sul decorso delle fibre mielinizzate si individuano precise sedi anatomo-funzionali: una “zona nodale” ove l’assone è spoglio di rivestimento, una “internodale” più o meno lunga e coperta da mielina e infine una di passaggio tra le due detta “iuxta-paranodale” [13]. Entro questa configurazione gli effettori della propagazione delle informazioni lungo la cellula nervosa sono i canali di membrana voltaggio-dipendenti. Li distinguiamo in rapidi e lenti a seconda del loro dominio temporale. I primi producono perturbazioni elettriche di membrana ad alto voltaggio e che esauriscono il loro ciclo nell’arco di circa 0,2-0,5 millisecondi (ms) mentre i canali lenti inducono perturbazioni di più basso voltaggio con cicli di esaurimento tra 0,5 e 10 ms. Negli assoni scarsamente mielinizzati e a bassa velocità di conduzione i canali sono diffusamente distribuiti entro la membrana e sono per lo più lenti. Negli assoni mielinizzati i canali rapidi tendono ad addensarsi nelle zone nodali mentre quelli lenti si diffondono con densità decrescente in senso iuxtaparanodale-internodale. Nella cellula nervosa c’è una notevole variabilità di distribuzione di diversi tipi di canali membranari: lungo l’assone prevalgono i canali voltaggio-dipendenti mentre nelle terminazioni dendritiche e nel soma il voltaggio della membrana è frutto principalmente di flussi ionici veicolati da canali attivati sia per via “endocrina” (per azione di neuromodulatori come ad esempio gli oppiodi), sia endocellulare e sinaptica [54]. Le relazioni funzionali tra l’azione dei canali rapidi e quella dei canali lenti sono alla base della modulazione dell’eccitabilità di membrana: la possibilità che in un punto della membrana possa verificarsi un potenziale elettrico è conseguenza del voltaggio che in quel punto è stato raggiunto in virtù di un potenziale precedente di una certa durata. Cruciale è la comprensione del risultato di dette interazioni: una sequenza di perturbazioni di membrana (portatrice della descrizione elettrica di un evento) viene recepita in un polo della cellula, confrontata con altre sequenze di perturbazioni secondo un algoritmo di addizioni-sottrazioni di voltaggio finché la perturbazione finale emergente sul soma cellulare non è trasformata in sequenze di potenziali sovramassimali (“spikes”) intercisi da intervalli variabili di silenzio elettrico. La distribuzione temporale dei “silenzi” (o, per definirla altrimenti, l’inverso della frequenza di scarica) corrisponde al grado di eccitabilità membranaria, ossia alla durata del “periodo refrattario” tra un potenziale e il successivo. Questi ultimi eventi elettrici costituiscono una sequenza informazionale, trasmessa in modulazione di frequenza, propagata ad alta fedeltà verso il polo assonale all’apice del quale avverrà la transduzione dal linguaggio nervoso codificato in frequenza nel flusso informazionale biochimico trasferito nelle vescicole sinaptiche secondo una ben nota proporzione quantica. Questi aspetti verranno ripresi e dettagliati nel capitolo dedicato a Biomembrane, Canali Ionici (CI) e Sistemi Molecolari di Segnalazione/Trasduzione/Trasporto (33) e nello spazio dedicato alla scoperta del “Campo di Disturbo” da parte dei fratelli Huneke (11b).
Considerando il numero di elementi che partecipano alla modulazione del prodotto informazionale finale abbiamo modo di formularci un’idea di quanta variabilità possa esprimersi nella relazione individuo/ambiente e quindi, inevitabilmente, nel rapporto salute/malattia. Una proporzione può salvarci dal naufragio nell’indeterminatezza: la soglia di eccitazione di un neurone è inversamente proporzionale alla velocità di conduzione del suo assone che, a sua volta, è direttamente proporzionale al dominio di risoluzione spazio-temporale a cui appartiene. Questo perché per tutte le citate variabili anatomo-funzionali nelle fibre a bassa velocità di conduzione il periodo refrattario è più lungo e quindi minore è la frequenza massima di scarica (o “firing”). Tale proporzione indica poi che la struttura gerarchica dei dominii spazio-temporali, costruita sul dispositivo di coordinazione-inibizione che ne garantisce l’adeguato comportamento nell’ambiente, corrisponde a una sovrapposizione di ranges (ossia di “spettri quantitativi”) di soglia, a loro volta espressione di ranges di firing. In anni recenti l’analisi matematica di alcuni bioritmi ha preso a indagare la distribuzione per spettri di frequenza delle loro oscillazioni nel dominio del tempo [50].
La via nervosa competente ai dominii spazio-temporali di bassa risoluzione è la più attigua ai fenomeni interattivi connessi alla produzione dell’infiammazione poiché è quella in cui il flusso nervoso d’informazione trova grande abbondanza di siti di integrazione/elaborazione con quello biochimico (come già abbiamo visto a proposito delle interazioni nutrizionali/contrattili). Anatomicamente possiamo riconoscere un compartimento del SN Periferico afferente, delimitato a valle dalla recettorialità cutaneo-mucosa e che raggiunge i gangli spinali dorsali tramite nervi sensitivi e misti. Da questo compartimento, col tramite dei rami comunicanti grigi, le stesse afferenze raggiungono le lamine del corno dorsale midollare. Qui si articola un reticolo nervoso multisinaptico in cui parte degli assoni converge sui nuclei intermedio-laterali midollari. Da tali nuclei numerosi assoni riemergono dal nevrasse per raggiungere i gangli pre- e laterovertebrali mentre altri vanno a costituire le vie multisinaptiche reticolari ascendenti e/o la via paleospinotalamica anteromediale per connettersi alle più alte stazioni bulbo-ponto-mesencefaliche e talamiche ipsi- e controlateralmente. Tramite i rami comunicanti bianchi (che entrano a far parte dei nervi spinali) emergono dai gangli pre- e laterovertebrali assoni efferenti che terminano nei plessi perivascolari cutanei e periviscerali a costituire il “Sistema Nervoso Autonomo Spinale”. L’architettura descritta è assai semplificata in quanto già tra gli assoni provenienti dai gangli autonomici e quelli provenienti dai plessi intramurali dei vasi ematici e linfatici esistono comunicazioni dirette (extraspinali) che danno luogo a fenomeni vasomotori da tempo noti come riflessi asso-assonici.
Il destino delle informazioni che, attraverso la liberazione di leve funzionali, scaturiscono dalle interazioni tra i flussi si rende evidente per lo più nei processi di centralizzazione nervosa delle vie polisinaptiche fino alle stazioni encefaliche sottocorticali. Meno chiara risulta l’elaborazione centralizzata del flusso nutrizionale-immunologico: qui vengono forse a prodursi competenze immunologiche inserite nella funzionalità di strutture altamente differenziate (linfociti ad altissima specificità recettoriale?).
30g.
Il caso della Compex Regional Pain Syndrome (CRPS): da fenomeno inusuale a paradigma
Fin qui abbiamo esaminato le modalità con cui il flusso informazionale nutrizionale-immunitario funge da sorgente di attivazione dell’attiguo e sovrastante flusso informazionale neuro-contrattile. Passiamo adesso alla descrizione di alcuni processi fisiopatologici orientati nella direzione opposta.
Tra le manifestazioni più interessanti sul piano clinico è quella che veniva definita “algodistrofia simpatico-riflessa”, entità nosografica recentemente rimaneggiata: se ne distingueva la variante “algodistrofia propriamente detta” e l’altra indicata come “causalgia”. L’elemento distintivo risiedeva nel fatto che nella seconda variante si poteva documentare un danno neuroperiferico attinente all’area somatica coinvolta. Dal 1994 il quadro viene definito “Complex Regional Pain Syndrome” (CRPS) ma nonostante gli sforzi di rinnovamento nosografico [7][18][49] esso poi ha finito per essere riclassificato in un Tipo I e in un Tipo II esattamente in base allo stesso criterio prima vigente: la presenza o meno di un danno neuroperiferico.
Nel Tipo I l’esordio può essere subdolo e variabile. Spesso il quadro clinico emerge dopo rimozione del tutore gessato resosi necessario per contenere una frattura o una dislocazione articolare ma talora compare inaspettatamente: ci si risveglia con la sindrome già attiva quando la sera precedente non si avvertiva alcun problema. Altre volte esso si instaura gradualmente per conclamarsi in pochi giorni o settimane. Spesso compare come abnorme manifestazione reattiva a traumi minori e, in tal caso, la caratteristica più eclatante è la sproporzione tra l’entità delle apparenti cause e quella degli effetti. Vale la pena di rammentare che alle sproporzioni tra le cause e gli effetti s’è già accennato a proposito delle soglie d’eccitabilità! Nel Tipo II, definito unicamente sul criterio della compresenza di un qualsiasi coinvolgimento nervoso loco-regionale rilevabile, l’accensione della sindrome si verifica ancor più subdolamente quando, in seguito a un evento traumatico da strappo, da scoppio o da taglio le previste fasi di riparazione tessutale deragliano lentamente verso un quadro clinico completamente diverso dalle aspettative. Le anamnesi remote talora rivelano un discreto novero di traumi pregressi diversi quanto a localizzazione, natura e gravità. Talvolta l’unico elemento che caratterizza questi pazienti, oltre al rilievo di un seppur lieve deficit neurologico concomitante, è una storia ricca di traumi (accidentali e/o chirurgici) ma non classificabili in base a relazioni anamnestiche codificate. In genere non vi sono dati circa precedenti eventi immunologici (malattie esantematiche o disimmuni) e la ricerca di indizi di patologia infettiva, immunitaria o metabolica non conduce ad alcun risultato significativo. Per quanto la CRPS tipo II si associ a una peggiore prognosi per la sussistenza di un danno nervoso le due varianti tendono nel tempo ad assomigliarsi soprattutto sul piano del potenziale rischio di invalidità residua e permanente [14].
Cerchiamo di individuare i costituenti del quadro clinico applicando un criterio sistematico: con il “dolore neuropatico” il SN esprime contemporaneamente un disordine delle sensibilità coscienti (costituito dalla triade allodinia, iperpatia e parestesia) e un disordine della motilità intenzionale e riflessa che generalmente è meno descritta. Pur senza sottrarre l’apporto necessario alla sopravvivenza tessutale il sistema vascolare esibisce fenomeni di disordine vasomotorio piuttosto caotico. Il sistema osteoarticolare mostra fenomeni di stampo infiammatorio con sovvertimento della matrice subcondrale, essudato sieroso articolare e osteoporosi delle estremità epifisarie. A quest’ultima è attribuita una tale importanza indiziaria per la diagnosi di CRPS da avere sollecitato la formulazione di neologismi radiologici che distinguono i fenomeni algodistrofici in tipo Modic I e Modic II [60] ove emergano immagini contemporaneamente indicative di fibrosi, edema e rarefazione ossea di uno o più segmenti osteoarticolari contigui. La distinzione tra tipo I e tipo II indica un passaggio di gravità del danno. I pochi resoconti clinici disponibili sugli esiti cronici convergono su condizioni di invalidità cronicizzata da anchilosi, distrofia dei tegumenti con maggiore vulnerabilità alle infezioni, atrofia muscolare da non uso, deformità articolari permanenti e dolore cronico ricorrente spesso poco rispondente agli antidolorifici. Fin qui gli aspetti clinici e fisiopatologici visti con l’approccio analitico della “malattia d’organo”. Se invece coniughiamo clinica e fisiopatologia nel rispetto della cifra di complessità che emerge da questa sindrome rileviamo quale primo elemento, anch’esso variabile e imprevedibile, la sua estensione topografica: possono risultare coinvolte uno o più dita, una mano o un piede, un intero arto e persino larga parte di un emisoma e tutto ciò senza alcuna apparente motivazione. L’elemento che invece rimane costante è il tipo di distribuzione topografica: se torniamo alla disposizione in “cavi” dei flussi informazionali lungo le vie anatomiche della periferia intra- ed extranervosa possiamo delimitare il disturbo in base a distribuzioni che riflettono diversi raggruppamenti di fibre su diversi livelli di gerarchia del nevrasse. In genere il neurologo clinico distingue patologie con distribuzione tronculare, plessurale e radicolo-metameriche fino a distribuzioni talamiche, extrapiramidali ed emisferiche. La distribuzione delle CRPS corrisponde invece alla topografia gangliare autonomico-spinale già descritta altrove (07) che ricalca un territorio diffuso che trasversalmente comprende tutti i flussi neuro/contrattili e nutrizionali/immunitari di un determinato distretto. Detti flussi informazionali vengono all’unisono rappresentati dall’alto di un piano elaborativo nervoso che ormai sappiamo essere alle prese soltanto con le espressioni “simboliche” dei suoi attori materiali. Sul successivo punto della nostra metodologia d’indagine (la dimensione temporale del fenomeno) alcuni autori hanno distinto la sindrome in una fase calda e in una fredda alludendo a un esordio di fenomeni infiammatori con evidenti manifestazioni vasomotorie seguito da una fase con prevalenza di rimaneggiamenti atrofico-cicatriziali post-lesionali. Entrambe le fasi sono state descritte in archi di tempo di settimane-mesi e la prognosi spontanea quoad valetudinem quantificata nell’arco di qualche anno è stata definita “mediamente infausta”. Nella nostra esperienza non abbiamo assistito a nette demarcazioni tra le due fasi ma abbiamo osservato che nell’ambito di una stessa CRPS circoscritta a un distretto possono contemporaneamente manifestarsi eventi riconducibili a una fase calda frammisti a quelli di una fase che potrebbe essere definita fredda. Probabilmente i processi alla base di questa patologia possono sovrapporsi a ondate di eventi ravvicinati e può risultare difficile demarcarne gli esiti nel tempo. Proviamo a individuare i punti d’incontro tra flussi informazionali, cioè l’ambito di elaborazione/integrazione da cui origina la perturbazione del bioritmo. L’esercizio interpretativo deve basarsi su una premessa affatto scontata: che ci si trovi di fronte a disordini scaturiti da un flusso neuro/contrattile anomalo al cospetto di un normale flusso nutrizionale/immunitario. Per quanto ne sappiamo questi pazienti sono esenti da movimenti perturbativi di tipo disimmune tanto che la stessa formulazione diagnostica di CRPS avviene una volta che sia escluso ogni preesistente coinvolgimento del Sistema Immunitario. Si tratta pertanto di una patologia che procede dall’alto: è il SN che accende l’infiammazione. Il concetto di flogosi neurogena è stato formulato da più di un secolo ma il suo accoglimento come fenomeno fisiopatologico di base costituisce una vicenda movimentata. Per una più estesa trattazione del tema è disponibile il capitolo (32) e i filmati (FEF1) (FEF2). Le stesse caratteristiche fisiopatologiche e cliniche rievocate in questa trattazione sono state già oggetto di studi appassionati in anni in cui l’esistenza delle citochine infiammatorie ancora non era nota e l’esplorazione dello stesso SN costituiva un’avventura eccitante quanto difficoltosa. In quegli anni gli studiosi sperimentarono la possibilità di suscitare diversi stati morbosi conseguenti a combinazioni di stimoli deboli con traumi localizzati imbattendosi così in stupefacenti fenomeni patologici che descrissero doviziosamente [47]. Ogni risultato sperimentale veniva registrato e fatto oggetto di analisi rispetto alle conoscenze acquisite permettendo ai successivi ricercatori di cercare prove per riordinare i risultati delle sperimentazioni precedenti secondo un modello teorico plausibile. Quella feconda esperienza venne purtroppo costretta in un cono d’ombra e quasi del tutto dimenticata allorché la ricerca biomolecolare iniziò a produrre evidenze dell’efficacia di alcune sostanze (antibiotici e FANS) contro malattie ad alto impatto sociale. Quasi tutte le risorse intellettuali e scientifiche si orientarono alla caccia della “molecola chiave” con cui malattie complesse e diverse potevano essere debellate. Le CRPS sono condizioni morbose poco responsive ai farmaci e pertanto è facile comprendere perché in quegli anni la ricerca del loro significato fisiopatologico (e di adeguate terapie) abbia sofferto di un grave rallentamento. L’interesse per la flogosi neurogena dovette sopportare un ulteriore periodo di oblio a favore della ricerca sulle malattie autoimmuni: con la scoperta degli anticorpi anticellulari (antinucleo, in particolare) si affermò un nuovo paradigma interpretativo di molte patologie croniche e poco altro si fece oltre che cercare l’autoanticorpo “chiave” ritenuto l’unico responsabile di malattie come la sarcoidosi e il neurolupus. I risultati di quel pervicace orientamento hanno fondato le strategie terapeutiche quasi esclusivamente sull’immunosoppressione. Solo quando quei furori si ridimensionarono ebbero modo di riemergere importanti contributi alla conoscenza della flogosi neurogena.
Sintetizzando le attuali concezioni fisiopatologiche sul tema azzardiamo una semplificazione circa l’interazione informazionale integrativo/elaborativa che esprime il quadro clinico delle CRPS. All’inizio si riteneva che i fenomeni fossero espressione di straordinarie sollecitazioni adrenergiche dei tessuti legate a eccesso di scarica nervosa (l’effetto di abnormi efferenze catecolminergiche da parte dei gangli simpatici pre- e laterovertebrali) mentre oggi per lo più ci si orienta verso la tesi di una forte sollecitazione, di uguale effetto, per eccesso di sensibilità recettoriale dei gangli spinali dorsali alle normali sollecitazioni catecolaminergiche [23]. Sta di fatto che una sindrome che fino a qualche tempo fa sembrava un “cigno nero” rispetto a tante entità nosologiche apparentemente intellegibili quanto a cause, manifestazioni cliniche e orientamento terapeutico viene oggi a rappresentare una costante sfida nella diagnosi differenziale. Trattandosi di patologie prive di markers biochimici diagnostici e in cui gli unici riscontri radiologici emergono solo quando la malattia ha già coinvolto lo stroma osteo-articolare è facile immaginare quanto gli usuali protocolli d’accertamento nei casi di dolore cronico tendano a risultare intempestivi (quando non omissivi) rispetto a una diagnosi di CRPS. Si segnala all’utente che ulteriori dettagli relativi alle CRPS sono illustrati in un capitolo dedicato alle Terapie con Anestetici Locali (TAL) nei disordini del trofismo tissutale (20).
30h.
Il percorso del flusso informazionale nervoso perturbato e la sua produzione di “solchi mnesici”
Abbiamo prima descritto una sindrome i cui fenomeni, se considerati singolarmente, non possono dirsi peculiari della CRPS. In altri termini la CRPS esibisce caratteristiche disfunzionali già organizzate e conglomerate autonomamente che tuttavia possiamo riconoscere isolatamente nel corso di varie altre sindromi. A proposito di malattie reumatologiche quando è stato menzionato lo stroma quale produttore fondamentale di leve funzionali perturbanti al piano mesenchimale abbiamo attribuito il ruolo di attore del fenomeno patologico piuttosto che quello del suo bersaglio. Nella CRPS lo stroma (derma, articolazioni, muscoli, connettivo osseo e vascolare) sembra invece costituire il bersaglio (assieme a strutture di altra derivazione embriologica come l’epidermide) se non altro perché non vi si riscontrano vettori della serie ematolinfocitaria normalmente implicati nei processi infiammatori (il cosidetto “infiltrato”). Tanto i fenomeni infiammatori quanto i loro esiti atrofico-fibro-cicatriziali sono innescati da leve funzionali immunitarie (torna la nota cascata citochinica TNF, IL2 e IL6) ma in una forma apparentemente bizzarra: poiché non si rinviene la primaria origine biochimica della noxa perturbatoria si può dire che nelle CRPS si esprime un effetto “senza sostanza”. In questa simbolizzazione riconosciamo l’origine nervosa della leva funzionale della sindrome ancora prima che in tanti altri dettagli. Anche sul piano degli esiti patologici tutte le manifestazioni lesionali della sindrome sono esercitate per via nervosa sostanzialmente per effetto delle disfunzioni vasomotorie indotte. Se ne può concludere che il flusso informazionale nervoso perturbato attiva da solo e dall’alto un flusso informazionale nutrizionale/immunitario sottostante senza che da questo provenga l’apporto della sua leva funzionale analogica (l’attivazione immunitaria di cellule, di vie del complemento e di immunoglobuline).
Nell’ambito di questo nostro modello interpretativo possiamo cogliere la caratteristica fondamentale dell’espressione patologica della CRPS: la “flogosi neurogena”. Complessivamente questi dati evocano l’idea di un repertorio comportamentale (cioè della manifestazione di un pattern predefinito di attivazione) e quindi può risultare illuminante individuare un corrispettivo ambientale in cui un simile pattern abbia un significato adattativo. Troviamo forti indizi nel comportamento di erbivori e carnivori ove un trauma localizzato di un arto induce rapidamente l’attivazione di un assetto funzionale di retrazione dell’arto offeso con la sua esclusione dalla funzione locomotoria. Questo schema non sembra direttamente correlabile all’entità del trauma né incidere gravemente sulla capacità motoria complessiva. In altri termini, quasi per non risultare d’intralcio nell’economia delle globali attività sensorimotorie, l’arto offeso sembra rispondere a una strategia adattativa globale attraverso l’autoesclusione. Più in basso nella scala filogenetica, tra i rettili e gli anfibi, la lesione dell’arto suscita dapprima la sua iniziale retrazione ma a questa segue rapidamente la autoelisione: in queste specie animali poco dopo la lesione l’arto diviene preda di forme rapidissime di atrofia a cui può fare seguito il completo distacco. Nell’arto in CRPS la ricerca della localizzazione lesionale con i criteri della semeiologia neurologica classica è frustrante: la lesione non presenta distribuzione tronculare, nè plessurale, nè radicolare. Piuttosto essa appare come un’anchilosi in retrazione in assenza di alcuna reale limitazione anatomica all’escursione articolare (almeno in fase precoce) e il dolore riflesso (che può rendere penoso persino toccare l’arto leso) aumenta la difficoltà dell’esame clinico diretto. Accettare che la flogosi neurogena sia espressione di un pattern contenuto in un “repertorio ancestrale” apre a diverse spiegazioni fisiopatologiche. Anzitutto la sindrome sembra serbare una forte relazione tra attivazione simpatica (caccia e competizione amorosa nell’animale, stress e traumatismo accidentale o chirurgico nell’uomo) e gestione neuromuscolare del movimento. Inoltre essa mostra un assetto sensorimotorio difensivo, retratto e ipersensibile. Infine si assiste a un tendenziale svincolo del microcircolo dalla regolazione circolatoria globale quasi che la periferia coinvolta si trasformasse quale organo a se stante anche sul piano nutrizionale. Questo modello interpretativo ci riconduce alla già menzionata strada del flusso informazionale neuro/contrattile che su questo livello di risoluzione spazio-temporale si avvale di “eventi simbolificati” e, pertanto, esclusivamente nervosi. Per definire un punto di inizio del flusso dobbiamo riferirci al polo afferente, ovvero al campo recettoriale periferico locoregionale: la molteplicità complanare e sovrapposta della risoluzione spazio-temporale qui si realizza attraverso la differenziazione del calibro e della mielinizzazione delle fibre (assieme alla loro diversa densità ed espansione topografica). Già sappiamo come la costituzione di ciascun bioritmo, corrispondente a ciascuna risoluzione spazio-temporale, si manifesti nel range di firing (ovvero nel processo di trasduzione analogico-digitale proveniente dal segnale ambientale esterno al soma del neurone afferente). Normalmente in questo processo il costante rapporto di trasduzione quantitativa tra evento analogico e frequenza di scarica sul soma del neurone garantisce la fedeltà qualitativa dell’informazione trasmessa. Tale rapporto, che rappresenta sul piano nervoso l’integrazione/elaborazione delle informazioni tra livelli di dominio spazio-temporale sovrapposti, è tuttavia soggetto a possibili alterazioni: se in un altro distretto, più o meno remoto ma comunque interconnesso lungo lo stesso flusso informazionale, si crea un sito di perturbazione (cioè un’altra interazione elaborativo/integrativa) può prodursi una leva funzionale che nella misura della sua capacità di attraversare quel flusso può alterare i rapporti quantitativi alla base della trasduzione analogico-digitale delle afferenze nervose. Gli studi neurofisiologici sulla sensitizzazione dell’afferenza somestesica sono stati moltiplicati dall’interesse clinico per il dolore neuropatico. Già si è accennato al fatto che questo tipo di dolore cronico è caratterizzato da tre fenomeni: l’allodinia (evocazione di una sensazione dolorosa in una modalità sensoriale diversa rispetto a quanto atteso dal tipo di stimolo), l’iperpatia (abbassamento della soglia di stimolo capace di evocare una sensazione dolorosa e sovvertimento delle sensibilità tattili-discriminative) e la parestesia (emergenza di una sensazione spontanea, non evocata da stimoli, molesta ma ben caratterizzata sul piano modale: calore, freddo, sensazioni puntorie o vibratorie, di contatto e di scorrimento di fluido sulla regione interessata). Nel dolore neuropatico sono state riconosciute le principali tappe di un processo fisiopatologico che è stato, per l’appunto, definito di “sensitizzazione”. I contenuti biomolecolari, elettrofisiologici e microscopici che sono emersi costituiscono una tale mole di dati che la loro trattazione sarebbe qui impossibile e dobbiamo limitarci a offrire poche indicazioni bibliografiche [34][36][59] e indugiare solo su alcuni concetti fondamentali. La sensitizzazione può avvenire per due vie convergenti: la prima è l’alterazione della distribuzione delle fibre nervose entro il campo recettoriale per perdita di sostanza secondaria a varie patologie tessutali (ad esempio nel diabete si manifesta quando avviene una prevalente distruzione di fibre amieliniche di piccolo calibro). L’altra via è quella della iperstimolazione dall’esterno tramite la reiterazione di microtraumi, ognuno dei quali di per sé incapace di alterare anatomicamente il tessuto. Già in questa duplicità patogenetica troviamo una forte consonanza con le due principali forme di CRPS: il Tipo II quale esito di lesione dell’innervazione loco-regionale e il Tipo I che può persino presentarsi privo di apparenti cause. Sul piano elettrofisiologico la sensitizzazione si manifesta con un fenomeno caratteristico: la stimolazione nervosa reiterata, a parità di intensità di stimolo, induce scariche di potenziali d’azione che aumentano progressivamente in frequenza aumentando la durata della stimolazione. In altri termini la reiterazione dello stesso stimolo produce da parte del neurone risposte in frequenza sempre più alte e questo vuol dire che si verifica un’amplificazione della propagazione delle informazioni attraverso la “modulazione in basso” della soglia di eccitabilità (FEF1). In termini neurofisiologici la sensitizzazione è un processo opposto all’accomodazione. Sul piano biomolecolare i responsabili di questo deragliamento del processo di trasduzione analogico-digitale sono i già citati canali ionici di membrana. La loro disposizione topografica, la loro costituzione chimica e la loro eterogeneità costituiscono i determinanti fondamentali dei processi di differenziazione filo-ontogenetica e dell’apprendimento nervoso. I fenomeni alla base di questa caratteristica adattativa risiedono nella modulazione dei Fattori di Crescita Embrionali e nella loro capacità di riattivarsi in seguito a sollecitazioni biochimiche provenienti da innumerevoli fonti tra cui, anzitutto, quelle indotte dai processi sintetici di livello sinaptico.
Comunque si realizzi la sensitizzazione induce una perturbazione nel flusso informazionale di appartenenza: l’incremento dell’eccitabilità esprime immediatamente un’elevazione di livello del dominio temporale a parità di estensione anatomica del reticolo in cui circolano le informazioni contenute in quel flusso. Questo autentico affollamento di informazioni pervade rapidamente tutta l’area di estensione del flusso informazionale e non va trascurato che l’incremento dell’eccitabilità corrisponde anche all’amplificazione della velocità di conduzione. Dal polo afferente (ove la sensitizzazione si è generata) al polo efferente (su cui il flusso si riversa) si espandono all’unisono gli effetti motori e secretori dell’attivazione efferente venendosi così a disarticolare gravemente le capacità coordinatorie-inibitorie dei dominii informazionali sovrastanti. Questi ultimi sono infatti tanto più ricchi di componenti inibitorie quanto più alto è il loro livello di sovrapposizione complanare gerarchica e quanto più segregate sono le vie in cui si muovono i flussi informazionali. La perdita dell’equilibrio tra il grado di risoluzione temporale (incrementata) e la sua estensione spaziale entro il proprio ambito reticolare (immutata) fa del flusso informazionale perturbato un fattore di propulsione del disordine “verso l’alto”. Non sono ancora note le “leve funzionali” attive in questo grado di elaborazione/integrazione tra flussi informazionali mentre sicuramente lo è il fenomeno neurofisiologico conseguente: sappiamo con certezza che la propagazione nel tempo e nello spazio della sensitizzazione, dalla superficie recettoriale afferente fino ai livelli metamerici spinali, elicita l’espansione del disordine mediante propagazione trans-sinaptica [57]. Questo dato corrobora anche qui il modello precedentemente proposto della trasmissione della perturbazione contemporanea su due tragitti, longitudinale e trasverso, come avviene in una miccia accesa. Se possiamo delineare la dimensione spazio-temporale del flusso informazionale della flogosi neurogena non è altrettanto facile comprenderne le conseguenze sul piano adattativo, individuale e di specie, né l’articolazione del fenomeno patologico nella sua cronicità. Per quanto riguarda il movente patogenetico postulato (la sproporzione tra dimensione temporale ed espansione spaziale del flusso nervoso) osserviamo che la conseguenza dell’accensione della perturbazione comporta nel lungo periodo il riassetto delle proporzioni: ai fenomeni destruenti provocati dalla flogosi neurogena fanno seguito fenomeni istologici che rimodellano profondamente la periferia innervata e che in questo sito sono illustrati più dettagliatamente altrove (32) (FEF1) (FEF2).
Le stesse “leve funzionali” vasomotorie, ghiandolari e muscolo-striate responsabili della fase calda della CRPS attivano per seconda intenzione quelle del flusso nutrizionale/immunitario che stimolano la guarigione per sostituzione fibroconnettivale dei tessuti specializzati. Ne risulta la perdita della funzionalità specifica loco-regionale ma allo stesso tempo si verifica la redistribuzione della recettorialità afferente su un livello ben più rarefatto: su un dominio spazio-temporale ancestrale. Il “trascinamento in basso” del dominio spazio-temporale della periferia sede della perturbazione metterà al bilancio quella stessa periferia in una condizione meno predisposta alla riaccensione della sensitizzazione e quel settore verrà pertanto a rappresentare (per il flusso di informazioni che viaggiano nel reticolo informazionale globale) un punto di alta soglia nel tragitto, una sorta di “dosso”. Un esempio mirabile di dosso è rappresentato da quei processi cicatriziali che esprimono capacità patogenetiche (come indicarono i fratelli Huneke). In quelle sedi si individua infatti il punto di demarcazione tra flussi informazionali differenti la cui transitoria soppressione funzionale mediante Anestetici Locali innesca una rimodulazione del traffico di informazioni. Volendo proporre una argomentazione finalistica: più che l’espressione di una semplice patologia i fenomeni di sensitizzazione sembrano rappresentare una modalità con cui l’esperienza traumatica individuale viene biologicamente trasformata in dispositivo adattativo. Guardando nell’insieme il processo per come lo abbiamo descritto sembra quasi che, al netto dell’episodio, il risultato sia la riformulazione dell’architettura strutturale e informazionale dei sistemi coinvolti, soprattutto di quelli posti sui livelli nervoso-centrali immediatamente attigui e sovrastanti. Qualunque sia il bilancio finalistico tale riformulazione diviene il nuovo terreno su cui viaggeranno, nel corso della vita di ogni individuo, le successive relazioni con l’ambiente: per ogni esito di patologia infiammatoria l’organismo offrirà all’ambiente una nuova mappa reticolare di soglie (ovvero ciò che abbiamo prima definito “solchi mnesici”) su cui potranno verificarsi, o meno, ulteriori processi patologici. Le tante possibili dissertazioni sugli aspetti epigenetici di tali informazioni non possono trovare spazio in questo capitolo.
30i.
Conclusioni e riferimenti tematici al sito
Da quanto esposto emergono chiaramente alcuni elementi tra loro strettamente connessi in una logica circolare. Anzitutto l’analisi dell’organizzazione dei sistemi viventi lascia intendere che essi interpretino come minacce per la loro integrità ogni forma di sovraccarico funzionale e informazionale ove le due modalità quasi sempre finiscono concretamente con il coincidere. E’ questa la logica che ha mosso l’evoluzione nella realizzazione di dispositivi di inibizione che risultano tanto più numerosi ed efficaci quanto più si prendano in esame organismi evoluti e complessi. L’inibizione di cui si parla è per lo più diretta a contenere l’espressione di dinamiche biologiche proprie: l’idea che essa sia prevalentemente orientata al contenimento di attività aggressive di nemici esterni è, a dir poco, preottocentesca. Lo stato di salute di un sistema vivente dipende largamente dall’efficacia dei dispositivi biologici di inibizione di cui è fornito e ciò è tanto più tangibile quanto più esso è evoluto e complesso. Problemi interpretativi possono derivare dal fatto che tale inibizione non è paragonabile a quella di un sasso che contrasta stolidamente un sistema inerziale sottostante quanto piuttosto a un sistema computazionale che si modella in architetture variabili nel tempo e nello spazio. Per facilitare la comprensione di questo concetto proponiamo una metafora: immaginiamo una cascata di vernici colorate che si abbatta su un pittore che, invece del pennello, disponga di un tampone per cancellare. Egli potrà asportare volumi di vernice così che dal magma multicolore possano emergere le forme, i colori desiderati e persino alcune illusioni di movimento.
Sappiamo che i sistemi inibitori sono parte integrante delle vie di comunicazione nei viventi. A questo punto possiamo pensare che se in tali vie di comunicazione si stabiliscono “aberrazioni” (ossia fenomeni che, in senso lato, inducano patologie) la possibilità di manipolare i sistemi inibitori possa incidere sull’andamento delle malattie. Questa risorsa è la base di quella terapia medica “patogenetica” che oggi è parte preponderante delle applicazioni farmacologiche a disposizione del medico. Se pensiamo ai vantaggi che cerchiamo dalla maggioranza dei presidi medici oggi utilizzati già quasi emerge la consapevolezza delle strutturazioni patologiche che abbiamo descritto: escludendo antibiotici e antineoplastici nella maggior parte dei casi somministriamo sostanze che mirano ad antagonizzare i dispositivi che l’organismo spontaneamente attiva nel corso delle patologie: antiinfiammatori (cioè inibitori dei principali mediatori del Sistema Immunitario aspecifico, il sistema delle prostaglandine e ancora altri inibitori delle citochine leucocitarie), anti-istaminici e altri anti-neuroimmunoregolatori che appartengono al nostro bagaglio biologico, calcio-antagonisti, alfa- e beta-bloccanti, anti-fosfodiesterasici, eccetera. Tale armamentario “soppressivo” è chiaramente frutto della reale relazione tra uomo e malattia e qualunque sia stato il motivo originario (empirico, biologico, bioingegneristico) per l’impiego di questi medicamenti essi hanno innegabilmente colto nel segno dimostrando di funzionare: infatti tra essi ci sono anche i nostri salva-vita. Ma qui torna il problema iniziale. Spesso siamo di fronte a un difetto che è nato “piccolo” e apparentemente banale e che ha tratto avvìo da una dinamica periferica e isolata. Lo sviluppo successivo della patologia è stato il frutto della messa in opera di meccanismi omeostatici che, per motivi intercorrenti e non riproducibili (si può stabilire un modo riproducibile con cui una pallina da golf si può perdere in un prato?) risale verso l’apice della gerarchia espandendosi al contempo su tutti i rivoli che trova disponibili. Quando tutto l’organismo è ormai preda dello squilibrio regolatorio il buon medico ne interpreta l’attore principale e lo bersaglia, spesso vincendo, scagliandogli contro il suo principale nemico rappresentato dal farmaco antagonista. Per esempio: contro l’attacco asmatico applichiamo il salbutamolo (agonista dell’inibizione sul broncospasmo e la secrezione mucosa) e il paziente torna a respirare. Se però ci ostiniamo a somministrarlo a chi non soffra di crisi asmatiche o all’asmatico che non sta manifestando crisi acute non proteggeremo il soggetto sano dal rischio di contrarre l’asma né l’asmatico dalla sua propensione a manifestare attacchi. In entrambi i casi provocheremmo piuttosto una malattia cronica dovuta dall’inopportuna somministrazione di salbutamolo. Per agire intelligentemente sfruttando l’inibizione dei sistemi regolatori informazionali dobbiamo quindi comprendere come essi si articolano nel tempo e nello spazio, ovvero entro quali ambiti si collocano nei ritmi biologici.
Siamo di fronte a un sistema che quando è “sano” tende ad assomigliare a un orologio meccanico le cui traiettorie informazionali sono riconducibili a quelle di una rete di veloci autostrade mentre quando è “malato” tende ad assomigliare a un magma uniforme e pervasivo le cui traiettorie informazionali si perdono in sentieri e tratturi lunghissimi, circonvoluti e inconcludenti. Abbiamo adesso gli elementi interpretativi per capire quello che è successo quando un sistema “si ammala”: è venuto meno il sistema inibitorio che delimita e costituisce, come uno scalpellino, la gerarchia adattativa strutturatasi nei tempi lunghissimi dell’evoluzione.
Abbiamo fin qui inteso fornire un’interpretazione di alcune conoscenze di Fisiologia e di Patologia già note e largamente condivise dalla comunità scientifica attenendoci a un modello allo stesso tempo evoluzionistico e biocibernetico. All’interno di questo modello le Terapie con Anestetici Locali (TAL) riescono a trovare spazio in termini di spiegabilità e coerenza scientifica laddove le comuni interpretazioni fisiopatologiche di “salute” e “malattia” sembrano restare ancora inchiodate alle relazioni lineari di causalità. E’ come se nel campo della Fisica non fossero mai avvenute le rivoluzioni delle teorie newtoniane rappresentate dalla relatività einsteiniana e dalla moderna quantistica. I frutti speculativi di questo nostro sforzo interpretativo possono essere così sintetizzati:
1) l’organizzazione funzionale dei viventi coincide con la loro “geometria dinamica”, in particolare con i movimenti di componenti mobili entro architetture che consentono scambi energetico-informazionali tra compartimenti ancestralmente più statici. In questa definizione di flussi tra interfacce abbiamo individuato due estremi di un ventaglio: da un lato la trasmissione elettrochimica propria del Sistema Nervoso, dall’altro la trasmissione metabolico-enzimatica nel tessuto interstiziale diffuso e comune a tutti i distretti dell’organismo. In questo dualismo è stato inserito il concetto di dominio informazionale spazio-temporale e delineato le relazioni dinamiche che intercorrono tra dominii adiacenti superando così la semplice continuità anatomica tra organi come unico presupposto per la propagazione di effetti patologici e terapeutici. Attraverso questo modello interpretativo abbiamo cercato di chiarire la possibilità di influire su effettori biologici distanti e apparentemente discontinui mediante un atto fortemente perturbativo, l’iniezione di Anestetici Locali (AL) inserito correttamente nella piramide dei dominii sovrapposti.
2) Analizzate le caratteristiche semi-quantitative dei tessuti viventi, ciascuno nel proprio dominio spazio-temporale, ne abbiamo apprezzato le principali qualità computazionali. Attraverso la nostra interpretazione dei processi di sensitizzazione-adattamento e di quelli legati alla deformazione plastica dei substrati possiamo ora spiegare perché una stessa dose di AL, iniettata nell’identico distretto somatico dello stesso individuo in momenti diversi del suo decorso clinico, possa evocare risposte diverse per intensità, durata e coinvolgimento di distretti remoti. Quest’esperienza ha messo in discussione due principali pilastri del “buon senso” che guida l’interpretazione clinica degli effetti terapeutici: il mantenimento di proporzionalità tra entità di perturbazione ed entità di risposta e la costanza dell’evocazione dell’effetto rispetto alla causa. Al di là dell’importanza degli eventi terapeutici empirici che si immaginava potessero sostenerla uno dei primi motivi della marginalizzazione da parte dell’Accademia del Novecento delle TAL, e ancor più della sua forma conosciuta come Terapia Neurale secondo Huneke (TNH), va cercato in questa apparente violazione del buon senso comune. Pur senza nutrire particolari aspirazioni a confliggere con la vigente metodologia evidence based pensiamo a quanto il suo rigoroso approccio statistico-matematico vacillerebbe davanti al dubbio di una relazione non più così costante tra dosi somministrate e gli attesi effetti clinico-strumentali.
3) Oltre che addurre evidenze fisiologiche già note, nella proposizione del nostro modello abbiamo indugiato sul concetto di “solco mnesico” che aspira a rappresentare con moderna concettualità la riedizione del termine “campo di disturbo” formulato dagli Huneke. Interpretando come evento dinamico e biocibernetico la produzione di un fenomeno di facilitazione informazionale (elettrochimica ed enzimatica) all’interno di un tessuto e considerando tale facilitazione quale esito strutturale di un’interazione perturbativa tra organismo e ambiente, l’applicazione terapeutica di AL “per certi versi” non può essere considerata alla stregua di un’interazione tra un principio attivo e un target cellulare secondo i canoni del modello farmacodinamico dell’effetto dose-dipendente. La durata dell’effetto farmacologico di un AL in basse concentrazioni e in relazione alla via di somministrazione presenta un range temporale di 30-40 minuti. Mentre le convenzionali terapie farmacologiche di patologie croniche si basano sulla reiterata assunzione di sostanze per mantenere i princìpi attivi a concentrazioni sieriche terapeutiche le TAL non confidano sulla durata del blocco della trasmissione nervosa e neppure sull’azione biochimica enzimatica su recettori nervosi o su mediatori delle cascate infiammatorie: si parla invece di “evento informazionale”. Così come la produzione dei solchi mnesici sono frutto di fenomeni patogenetici altrettanto informazionali (come la reiterazione di eventi perturbativi, il coinvolgimento contemporaneo o consecutivo di flussi di dominio spazio-temporali adiacenti, eccetera) il blocco della conduzione elettrochimica rappresenta un evento, un episodio. La caratteristica di tale evento, assai raro in natura a parte la folgorazione elettrica, è quella di porre un segno “meno”, un’interruzione entro un sistema computazionale. Così l’eventuale effetto a distanza non è il frutto diretto dell’evento ma la registrazione mnesica di un segno algebrico di verso negativo nella definizione del calcolo finale che tendenzialmente verrà mantenuto a lunga distanza di tempo. Tale effetto potrà mostrare maggiore efficacia se inserito nella sede più spiccatamente computazionale dell’organismo: il Sistema Nervoso. Questi stessi concetti, scientificamente convalidati e universalmente apprezzati, sono alla base della persistente efficacia della terapia elettroconvulsivante nell’epilessia e nel disturbo depressivo maggiore così pure della cardioversione elettrica in caso di fibrillazione ventricolare. D’altra parte tale proprietà informazionale del resetting, con le sue possibilità applicative in terapia, non costituisce esclusivo appannaggio del livello informazionale neuro/contrattile. Nella dismenorrea, ad esempio, è sfruttata da tempo l’efficacia terapeutica dei farmaci anticoncezionali prescritti per pochi mesi: nell’ambito di questi bioritmi più lenti a cosa altro potrebbe corrispondere il razionale dell’effetto degli estro-progestinici imposti artificialmente entro il ciclo mestruale se non nel meccanismo del blocco/reset?
Per altri versi l’efficacia terapeutica degli AL si esprime anche grazie all’affinità che mostrano nei confronti di proteine che assumono ruoli cruciali nelle segnalazioni intracellulari, intercellulari e tra cellule e ambiente extracellulare in entrambi i sensi di marcia. Questa nozione, oggi sempre più corroborata e pregna di ricadute sul piano clinico, trasse origine da studi iniziati all’alba di questo Secolo sui recettori muscarinici M1 ricombinanti espressi negli ovociti che identificarono per gli AL un sito d’azione intracellulare comune su diversi recettori accoppiati alla proteina “G” (33) (34) (35).
4) Il presente testo nasce inizialmente dalla volontà di rimarcare alcune relazioni anatomiche e funzionali tra i sistemi Nervoso, Immunitario ed Endocrino. L’attuale tendenza a unificare queste tre entità, la cui parentela reciproca emerge proprio dai comuni aspetti biocibernetici, ha partorito diverse proposte di modello dottrinario. Alcune di esse risultano influenzate da derive metafisiche mentre altre sono più attinenti a una moderna concezione scientifica materialistica. Per quanto riguarda la neuro-immunomodulazione mediante AL il nostro contributo alla comprensione dei suddetti sistemi computazionali coincide con quanto avevano già evidenziato le esperienze storiche sulla TNH e, successivamente, le nostre attuali esperienze. Nel modello qui proposto è la disposizione gerarchica dei domini spazio-temporali che permette l’orientamento tra i diversi effettori (nervosi, immunitari e ormonali) rendendo quasi intuitiva e prevedibile la relazione tra la patologia e le pratiche cliniche di neuro-immunomodulazione quali sono le TAL.
[1] Andreev NY, Dimitrieva N, Koltzenburg M, McMahon SB
Peripheral administration of nerve growth factor in the adult rat produces a thermal hyperalgesia that requires the presence of sympathetic post-ganglionic neurons
Pain, 63(1):109-15, Oct 1995
[2] Ascher NL, Hoffman RA, Hanto DW, Simmons RL
Cellular basis of allograft rejection
Immunol Rev, 77:217-32, 1984
[3] Baglan H, Lazzari C, Guerrieri F
Learning in mosquito larvae (Aedes aegypti): Habituation to a visual danger signal
Jour Insect Physiol. Apr;98:160-166, 2017
[4] Bershteyn M, Nowakowski TJ, Pollen AA, Di Lullo E, Nene A, Wynshaw-Boris A, Kriegstein AR
Human iPSC-Derived Cerebral Organoids Model Cellular Features of Lissencephaly and Reveal Prolonged Mitosis of Outer Radial Glia
Cell Stem Cell, 20(4):435-449, 6 Apr 2017
[5] Blalock JE
A molecular basis for bidirectional communication between the immune and neuroendocrine systems
Physiol Rev, 69(1):1-32, Jan 1989
[6] Blatteis CM
Fever: Pathological or physiological, injurious or beneficial?
Jour of Thermal Biolog, 28(1):1-13, January 2003
[7] Bruehl S, Harden RN, Galer BS, Saltz S, Bertram M, Backonja M et Al
External validation of IASP diagnostic criteria for complex regional pain syndrome and proposed research diagnostic criteria.
Pain, 81(1-2):147-54, May 1999
[8] Carbone A, Gromov M, Przemsyl P
Pattern Formation in Biology, Vision and Dynamics Wordl Scientific, 2000
[9] Carlson NG, Wieggel WA, Chen J, Bacchi A, Rogers SW, Gahring LC
Inflammatory cytokines IL-1 alpha, IL-1 beta, IL-6, and TNF-alpha impart neuroprotection to an excitotoxin through distinct pathways
Jour Immunol, 163(7):3963-8, Oct 1999
[10] Chaplin DC
Overview of the immune response
Jour Allergy Clin Immunol, 125(2 Suppl 2):S3-23, Feb 2010
[11] Davidsen K, Matsen FA 4th
Benchmarking Tree and Ancestral Sequence Inference for B-Cell Receptor Sequences
Front Immunol, 9:2451, Oct 2018
[12] De Blecourt J, Polman A, De Blecourt-Eindersma
Hereditary factors in rheumatoid arthritis and ankylosing spondylitis
Ann Rheum Dis, 20:215-20, 1961
[13] Debanne D, Campanac E, Bialowas A, Carlier E, Alcaraz G
Axon physiology
Physiol Rev, 91(2):555-602, Apr 2011
[14] Dey S, Guthmiller KB, Varacallo M
Complex Regional Pain Syndrome
In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing, Jan 2022
[15] Eccles JC
The Inhibitory pathways of the central nervous system
Liverpool Univ Press, 1969
[16] Freidin M, Bennett MV, Kessler JA
Cultured sympathetic neurons synthesize and release the cytokine interleukin beta
Proc Natl Acad Sci, 89(21):10440-3, nov 1992
[17] Fries JF, Hochberg MC, Medsger TA Jr, Hunder GG, Bombardier C
Criteria for rheumatic disease. Different types and different functions.
The American College of Rheumatology Diagnostic and Therapeutic Criteria Committee
Arthritis Rheum, 37(4):454-62, Apr 1994
[18] Harden RN, Bruehl S, Galer BS, Saltz S, Bertram M, Backonja M et Al
Complex Regional Pain Syndrome: are the IASP diagnostic criteria valid and sufficiently comprehensive?
Pain, 83(2):211-9, Nov 1999
[19] Hasler P, Zouali M
B cell receptor signaling and autoimmunity
FASEB Jour, 15(12):2085-98, Oct 2001
[20] Hill M, Goldspink G
Expression and splicing of the insulin-like growth factor gene in rodent muscle is associated with muscle satellite (stem) cell activation following local tissue damage
Jour Physiol, 549(Pt 2):409-18, Jun 2003
[21] Hirth M, Rukwied R, Gromann A, Turnquist B, Weinkauf B et Al
Nerve growth factor induces sensitization of nociceptors without evidence for increased intraepidermal nerve fiber density
Pain, 154(11):2500-2511, Nov 2013
[22] Hollmann MW, Wieczorek KS, Berger A et Al
Local anesthetic inhibition of G protein‐coupled receptor signaling by interference with Galpha(q) protein function
Mol Pharmacol, 59: 294-301, 2001
[23] Jänig W, Levine JD, Michaelis M
Interactions of sympathetic and primary afferent neurons following nerve injury and tissue trauma
Prog Brain Res, 113:161-84, 1996
[24] Jerne NK
Towards a network theory of the immune system
Annales d’immunologie, 125C (1-2): 373-389, 1974
[25] Julio-Pieper M, Flor PJ, Dinan TG, Cryan JF
Exciting times beyond the brain: metabotropic glutamate receptors in peripheral and non-neural tissues
Pharmacol Rev, 63(1):35-58, Mar 2011
[26] Koch-Henriksen N, Sorensen PS
The changing demographic pattern of multiple sclerosis epidemiology
Lancet Neurol, 9(5):520-32, May 2010
[27] Kohm AP, Sanders VM
Norepinephrine and beta 2-adrenergic receptor stimulation regulate CD4+ T and B lymphocyte function in vitro and in vivo
Pharmacol Rev, 53(4):487-525, Dec 2001
[28] Leon A, Buriani A, Dal Toso R, Fabris M, Romanello S, Aloe L et Al
Mast Cells synthesize, store, and release Nerve Growth Factor
Proc Natl Acad Sci, 26;91(9):3739-43, Apr 1994
[29] Levi-Montalcini R, Skaper SD, Dal Toso R, Petrelli L, Leon A
Nerve growth factor: from neurotrophin to neurokine
Trends Neurosci, 19(11):514-20, Nov 1996
[30] Lippi G, Guidi G
Lipoprotein(a): from ancestral benefit to modern pathogen?
QJM, 93, 2, 75-84, 2000
[31] Maes M
Evidence for an immune response in major depression: a review and hypothesis
Prog Neuropsychopharmacol Biol Psych, 19(1):11-38, Jan 1995
[32] Marti HJ, Bernaudin M, Bellail A, Schoch H, Euler M, Petit E et Al
Hypoxia-induced vascular endothelial growth factor expression precedes neovascularization after cerebral ischemia
Am Jour Pathol, 156(3):965-76, Mar 2000
[33] Matsuoka I, Ohkubo S
ATP- and adenosine-mediated signaling in the central nervous system: adenosine receptor activation by ATP through rapid and localized generation of adenosine by ecto-nucleotidases
Jour Pharmacol Sci, 94(2):95-9, Feb 2004
[34] Millan MJ
The induction of pain: an integrative review
Prog Neurobiol, 57(1):1-164, Jan 1999
[35] Mills JH, Kim DG, Krenz A, Chen JF, Bynoe MS
A2A adenosine receptor signaling in lymphocytes and the central nervous system regulates inflammation during experimental autoimmune encephalomyelitis
Jour Immunol, 188(11):5713-22, 2012
[36] Mizumura K
Peripheral mechanism of hyperalgesia-sensitization of nociceptors
Nagoya Jour Med Sci, 60(3-4):69-87, Nov 1997
[37] Molina H
Complement and immunity
Rheum Dis Clin North Am, 30(1):1-18, Feb 2004
[38] Nicol GD, Vasko MR
Unraveling the story of NGF-mediated sensitization of nociceptive sensory neurons: ON or OFF the trks?
Mol Interv, 7(1):26-41, Feb 2007
[39] Okajima A, Miyazawa K, Naitoh Y, Inoue K, Kitamura N
Induction of hepatocyte growth factor activator messenger RNA in the liver following tissue injury and acute inflammation
Hepatology, 25(1):97-102, Jan 1997
[40] Ouallet JC, Radat F, Creange A, Abdellaoui M, Heinzlef O, Giannesini C, Hautecoeur P et Al
Evaluation of emotional disorders before and during treatment with interferon beta in patients with multiple sclerosis
Jour Neurol Sci, 19;413:116739, Feb 2020
[41] Pecht I
Immuno-receptors: from recognition to signaling and function
Eur Biophys Jour, 47(4):363-371, May 2018
[42] Pierpaoli W, Sorkin E
Relationship between Thymus and Hypophysis
Nature, vol 215, 834-837, Aug 1967
[43] Ricker G
Pathophysiologie als Naturwissenschaft
Berlin, 1924
[44] Rosati G
The prevalence of multiple sclerosis in the world: an update
Neurol Sci, 22(2):117-39, Apr 2001
[45] Schnakenberg M, Thomas C, Schmelz M, Rukwied R
Nerve growth factor sensitizes nociceptors to C-fibre selective supra-threshold electrical stimuli in human skin
Eur Jour Pain, 25(2):385-397, Feb 2021
[46] Shweiki D, Itin A, Soffer D, Keshet E
Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis
Nature, 359(6398):843-5, Oct 1992
[47] Speransky AD
Grundlagen der Theorie der Medizin
Leningrad 1934 (Übersetzung der 3 Aufl von KR Roques, Berlin 1950)
[48] Stanton GJ, Weigent DA, Fleischmann WR Jr, Dianzani F, Baron S
Interferon review
Invest Radiol, 22(3):259-73, Mar 1987
[49] Stanton-Hicks M, Janig W, Hassenbusch S, Haddox JD, Boas R, Wilson P
Reflex sympathetic dystrophy: changing concepts and taxonomy
Pain, 63(1):127-33, Oct 1995
[50] Stefanovska A, Bracic M, Kvernmo HD
Wavelet analysis of oscillations in the peripheral blood circulation measured by laser doppler technique
IEEE Trans Biomed Eng, 46(10):1230-9, Oct 1999
[51] Taussig MJ
Molecular genetics of immunoglobulins
Immunol Suppl, 1:7-15, 1988
[52] Teuscher C, Subramanian M, Noubade R, Gao JF, Offner H, Zachary JF
Central histamine H3 receptor signaling negatively regulates susceptibility to autoimmune inflammatory disease of the CNS
Proc Natl Acad Sci, 104(24):10146-51, Jun 2007
[53] Thorn CE, Kyte H, Slaff DW, Shore AC
An association between vasomotion and oxygen extraction
Am Jour Physiol Heart Circ Physiol, 301(2):H442-9, Aug 2011
[54] Toescu EC, Verkhratsky A
Integrative aspects of calcium signalling
Jour Neurophysiol, 79, 240-252, 1998
[55] van Buul-Offers SC, R Kooijman R
The role of growth hormone and insulin-like growth factors in the immune system
Cell Mol Life Sci, 29;54(10):1083-1094, Jan 2014
[56] Wang X, Cao L, Liu S, Zhou Y, Jiarui Zhou J e Al
The critical roles of IGFs in immune modulation and inflammation
Cytokine, 183:156750, Nov 2024
[57] Watkins LR, Maier SF
Beyond neurons: Evidence that immune and glial cells contribute to pathological pain states
Physiol Rev, 82(4):981-1011, Oct 2002
[58] Wichers MC, Maes M
The role of indoleamine 2,3-dioxygenase (IDO) in the pathophysiology of interferon-alpha-induced depression
Jour Psychiatry Neurosci, 29(1):11-7, Jan 2004
[59] Woolf CJ
Central sensitization: Implications for the diagnosis and treatment of pain
Pain, 152(3 Suppl):S2-15, Mar 2011
[60] Zhang YH, Zhao CQ, Jiang LS, Chen XD, Dai LY
Modic changes: a systematic review of the literature
Eur Spine Jour, 17(10):1289-99, Oct 2008